 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Chilean Journal of Agricultural Research, Vol. 71, No. 2, April-June, 2011, pp. 205-211 RESEARCH Alternative Management Of A Problematic Weed Of Wheat Avena fatua L. by Metabolites of Trichoderma Alternativas de manejo de una maleza problemática detrigo Avena fatua L. por metabolitos de Trichoderma. Arshad Javaid1*, and Sajjad Ali1 1 Punjab University Lahore, Institute of Agricultural Sciences,
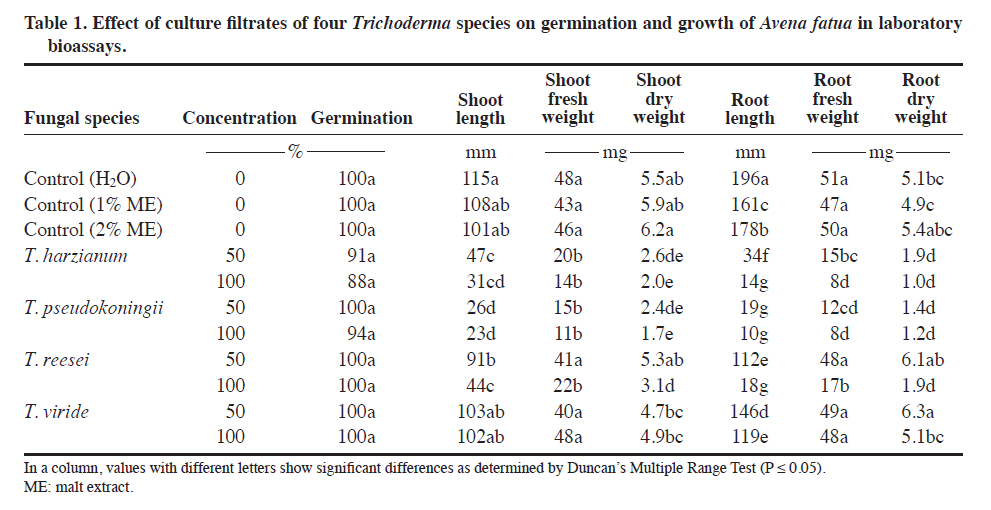
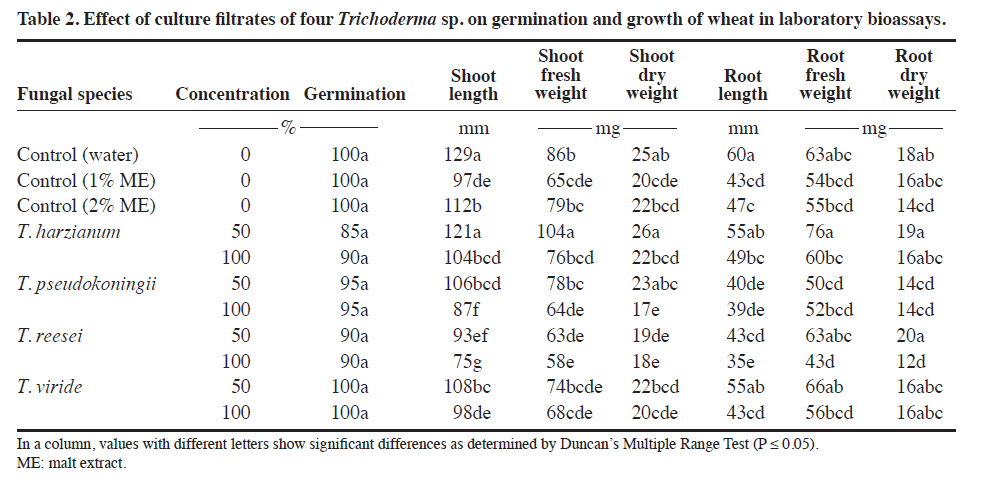
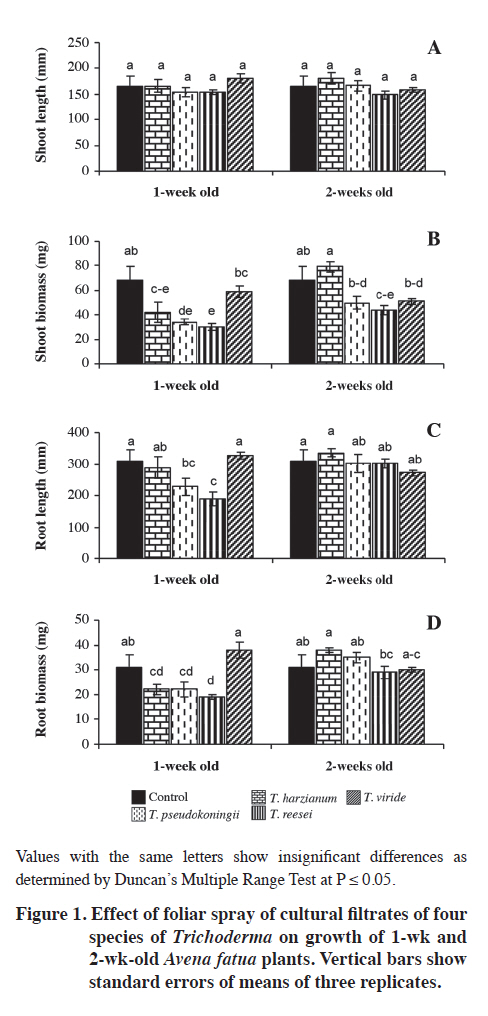
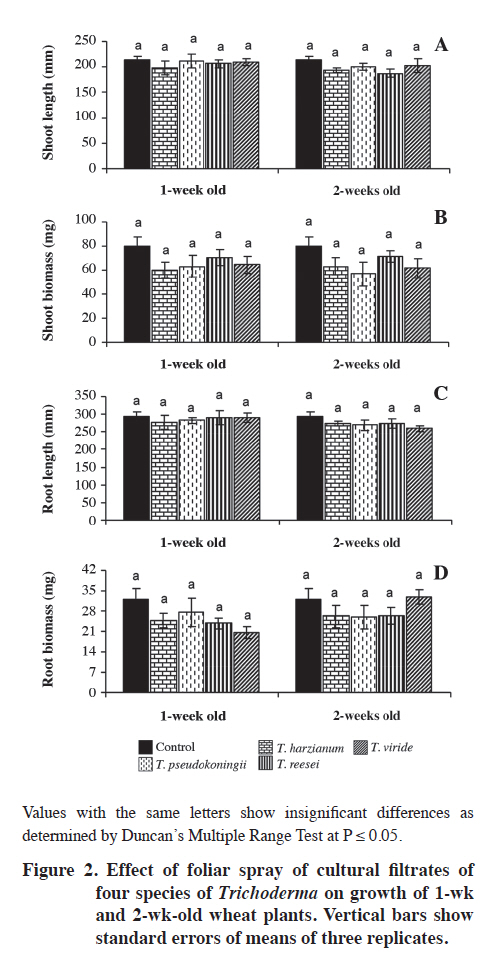
Lahore, Pakistan. Received: 8 October 2010. Code Number: cj11024 ABSTRACT Wild oat (Avena fatua L.) is a problematic weed of wheat (Triticum aestivum L.) in Pakistan. The present study was designed to evaluate the herbicidal activity of culture filtrates of four Trichoderma spp., namely T. harzianum, T. pseudokoningii, T. reesei and T. viride, against this weed species. In a laboratory bioassay, original (100%) and diluted (50%) culture filtrates of T. harzianum and T. pseudokoningii significantly reduced shoot and root growth of A. fatua seedlings. Only original culture filtrates of T. reesei exhibited significant effects, while the effect of filtrates of T. viride was insignificant against shoot and root growth of the target weed species. Generally, original concentrations of culture filtrates of all Trichoderma spp., except T. harzianum, significantly reduced various parameters of root and shoot growth of wheat seedlings. In foliar spray bioassay, pot-grown 1-wk and 2-wk old A. fatua and wheat seedlings were sprayed with 100% culture filtrates of the four Trichoderma spp. thrice with 5 d interval each. Culture filtrates of all except T. viride significantly diminished root and shoot biomass of A. fatua plants in 1-wk old plants. The effect of foliar spray on root and shoot growth of wheat was insignificant. Present study concludes that metabolites of T. harzianum, T. reesei and T. pseudokoningii contain herbicidal constituents for the management of A. fatua. Key words: Alternative herbicides, Avena fatua, fungal metabolites, Trichoderma spp. RESUMEN La avena silvestre (Avena fatua L.) es una maleza problemáticadel trigo (Triticum aestivum L.) en Paquistán. El presente estudio se diseñó para evaluar la actividad de filtrados de cultivos de cuatro Trichoderma spp.: T. harzianum, T. pseudokoningii, T. reesei y T. viride, contra esta especie. En un bioensayo de laboratorio, filtrados de cultivos originales (100%) y diluidos (50%) de T. harzianum and T. pseudokoningii redujeron significativamente elcrecimiento de brotes y raíces de plántulas de A. fatua. Sólo los filtrados de cultivos originales de T. reesei exhibieron efecto significativo mientras el efecto de filtrados de T. viride fue insignificante contra el crecimiento de brotes y raíces de la maleza. Las concentraciones originalesde filtrados de todos los cultivos de Trichoderma spp. excepto T. harzianum redujeron significativamente variosparámetros de crecimiento de brotes y raíces de plántulasde trigo. En el bioensayo de aspersión foliar, plántulasde A. fatua y trigo de 1 y 2 semanas de edad cultivadas en macetas se asperjaron con filtrados de cultivo 100% de las cuatro especies de Trichoderma tres veces con 5 d de intervalo cada uno. Los filtrados de cultivo de todos excepto T. viride redujeron significativamente la biomasade raíces y brotes de plantas de A. fatua en plantas de 1semana de edad. El efecto de la aspersión foliar sobre el crecimiento de raíces y brotes de trigo fue insignificante. El presente estudio concluye que los metabolitos de T. harzianum, T. reesei y T. pseudokoningii contienen constituyentes herbicidas para el manejo de A. fatua. Palabras clave: herbicidas alternativos, Avena fatua, metabolitos fúngicos, Trichoderma spp. In the literature, there are many examples of fungal products having herbicidal activities. AAl-toxin is a phytotoxin produced by Alternaria alternata (Fr.) Keissl. pathovars that infect tomato. Its mode of infection was reported to be inhibition of RNA synthesis. It is active on many weed species (Abbas et al., 1995). Two phytotoxic metabolites prehelminthosporal and dihydropore, were isolated from the cultural filtrates of the fungus Bipolaris sp. showing herbicidal activity against Sorghum halepense (L.) Pers. (Parmar and Devkumar, 1993). Three herbicidal metabolites, 3-hydroxybenzylalcohol, 2-methyl hydroquinone and epiepoformin were isolated from a soil-borne fungus Scopulariopsis brumptii. Similarly, a phytotoxic metabolite trans-4 aminoproline isolated from culture filtrates of Ascochyta caulina was found to be very effective in controlling Chenopodium album (L.) (Evidente et al., 2000). Evidente et al. (2005) isolated a new phytotoxic trisubstituted naphthofuroazepinone from the culture filtrates of Drechslera siccans, named drazepinone, and characterized as a 3,5,12 a-trimethyl- 2,5,5a,12a-tetrahydro-1H-naphtho[2′,3′:4,5]furo[2,3-b] azepin-2-one. Assayed at 2 μg μL−1 solution, the novel metabolite proved to have broad-spectrum herbicidal properties. Phyllosticta cirsii, a fungal pathogen isolated from diseased Cirsium arvense leaves and evaluated as a biocontrol agent of this noxious perennial weed, produces different phytotoxic metabolites with potential herbicidal activity when grown in liquid cultures. Phyllostictines A−D, four novel oxazatricycloalkenones, were isolated from this pathogen (Evidente et al., 2008). Recently Javaid and Adrees (2009) reported that metabolites of Alternaria alternata, Fusarium oxysporum, F. solani, Drechslera hawaiiensis, D. australiensis and D. rostrata were highly effective in controlling the growth of the noxious weed Parthenium hysterophorus. Trichoderma spp. are among the most frequently isolated soil fungi and present in plant root ecosystems (Harman et al., 2004). To date, most of the studies on Trichoderma spp. have been conducted with respect to their activity as biological control agents. Their efficacy has been demonstrated for the management of nematode and fungal diseases under field conditions (Sahebani and Hadavi, 2008; Hanada et al., 2009). The modes of action as biological control agents include mycoparasitism, antibiosis, competition, enzyme activity and induced plant defense (Howell, 2003). In addition to their role as biocontrol agents, Trichoderma spp. have also been used in a wide range of commercial enzyme productions, namely, cellulases, hemicellulases, proteases, and β-1,3 glucanase (Sadhasivam et al., 2008; Kovacs et al., 2009).However, weed control by using Trichoderma spp. is still a relatively unexplored field and the few known studies in this area are restricted to T. virens only (Hutchinson,1999; Heraux et al., 2005). The present study was, therefore, designed to evaluate the herbicidal activity of culture filtrates of four Trichoderma spp.: T. harzianum, T. pseudokoningii, T. reesei and T. viride, against Avena fatua, a problematic weed of wheat. MATERIALS AND METHODS Procurement of Trichoderma species Pure cultures of four species of Trichoderma: T. harzianum, T. pseudokoningii, T. reesei and T. viride were procured from the Fungal Culture Bank of the Institute of Plant Pathology, University of the Punjab, Lahore, Pakistan. These cultures were maintained on Malt Extract Agar (MEA) medium and stored at 4 °C in a refrigerator. Preparation of cultural filtrates of the test fungi Malt extract broth medium was autoclaved at 121 ºC in 250 mL conical flasks at 100 mL medium in each flask. Flasks were inoculated with 5 mm agar discs of each of the four test fungi from margins of actively growing fungal colonies. Inoculated flasks were incubated under static conditions at 25 °C for 4 wk. After 4 wk, the cultures were filtered through sterilized muslin cloth followed by Whatman filter paper N° 1. These filtrates were preserved at 4 °C in a refrigerator. The cultural filtrates were used within 1 wk of filtration to avoid any contamination or chemical alteration (Javaid and Adrees, 2009). Laboratory bioassay Mature seeds of A. fatua were collected from wheat fields of the University of the Punjab, Quaid-e-Azam Campus Lahore, Pakistan. Seeds were dried in the sun, packed in paper bags and stored at room temperature. Ten seeds of A. fatua and wheat var. Sehr were placed in 9 cm diameter sterilized Petri plates with sterilized filter papers moistened with 3 mL of original (100%) and diluted (50%) crude fungal extract of different Trichoderma species. The negative control treatment received 3 mL of sterilized distilled water. Since fungal metabolites were obtained using 2% malt extract broth, and filtrates were also used as 50% dilutions. Therefore, two positive control treatments with 3 mL of 2% and 3 mL of 1% malt extract broth were also included. Each treatment was replicated four times. The Petri plates were arranged in a completely randomized design in a growth room maintained at 18 ± 2 °C and 10:14 h photoperiod. Plants were harvested after 12 d. Data regarding germination, and root/shoot growth in terms of length and fresh biomass were recorded. Plants were dried at 60 ºC to constant weight and dry weight was recorded (Javaid and Adrees, 2009). Foliar spray bioassay Experiment was conducted in February-March 2009 at the University of the Punjab, Lahore (31º57’N;74º31’ E) Pakistan. Plastic pots of 8 cm diameter and 12 cm deep were filled with 350 g of sandy loam soil with 0.69% organic matter, pH 7.8, N 0.035%, available P 6.3 mg kg-1 and available K 100 mg kg-1. The micronutrients B, Mn, Fe, Cu and Zn were 1.06, 22.8, 10.8, 1.9 and 1.3 mg kg-1 of soil, respectively. Ten seeds of A. fatua and wheat var. Sehr were sown in each pot. After germination, pots were divided into two sets to perform the foliar spray on 1-wk and 2-wk old seedlings. All the pots were arranged in a completely randomized design under natural environmental conditions. Original culture filtrates of the four selected Trichoderma species were sprayed on 1-wk and 2-wk old A. fatua and wheat seedlings. Both of the sets were sprayed 3-times with an interval of 5 d. Treatment in a similar manner with distilled water spray served as control. All the plants were harvested after 40 d growth. Plants were carefully uprooted and washed under tap water. Roots were separated from shoots. Data regarding length and dry biomass of shoot and root were recorded. Statistical analysis All the data were subjected to ANOVA followed by Duncan’s Multiple Range Test (P ≤ 0.05) to delineate mean differences (Steel and Torrie, 1980). RESULTS Laboratory bioassay The effect of culture filtrates of all the Trichoderma species was insignificant on germination of A. fatua seeds (Table 1). Maximum shoot length of 115 mm was recorded in negative control (water). Decreases in shoot length in 1% and 2% malt extract treatments were insignificant. Culture filtrates of all the Trichoderma species, except T. viride, significantly reduced shoot length. Highest toxicity was recorded due to the culture filtrate of T. pseudokoningii, followed by T. harzianum and T. reesei, respectively. The original culture filtrates (100%) of T. pseudokoningii, T. harzianum, T. reesei and T. viride reduced the shoot length of A. fatua by 80, 73, 62 and 11%, respectively; as compared to the negative control (Table 1). Culture filtrates of 50%, as well as 100%, of T. pseudokoningii and T. harzianum significantly suppressed shoot fresh and dry biomass of A. fatua as compared to negative control. Similarly, the negative effect of 100% filtrates of T. reesei was significant on shoot fresh and dry biomass. Culture filtrates of T. viride, however, failed to exhibit any pronounced effect on shoot biomass of A. fatua (Table 1). Root length was reduced due to 1% and 2% malt extract by 18 and 9%, respectively. Culture filtrates of all four tested Trichoderma species significantly reduced root length. The adverse effects of T. pseudokoningii and T. harzianum filtrates on root length were more pronounced than the effects of T. reesei and T. viride. There was 95, 93, 91 and 39% reduction in root length due to 100% culture filtrates of T. pseudokoningii, T. harzianum, T. reesei and T. viride, respectively, as compared to negative control. Malt extract medium exhibited insignificant effects on root fresh and dry biomass. The effect of culture filtrates of the four Trichoderma species on fresh and dry biomass of roots of A. fatua was generally similar to that observed on root length (Table 1). None of the treatments exhibited significant effects on germination of wheat seeds. Except for 50% of culture filtrates of T. harzianum, all treatments significantly reduced shoots length as compared to negative control. Similarly, original culture filtrates (100%) of all the Trichoderma spp. except T. harzianum significantly reduced shoot biomass of wheat seedlings. The effects of culture filtrates of the different Trichoderma species evaluated were in general similar for both root and shoot growth parameters (Table 2). Foliar spray bioassay Even though a minimum effect of the different Trichoderma extracts was observed on the shoot length of A. fatua, there was a clear suppressing effect on shoot biomass of the extracts when they were sprayed 1 wk after germination. The effect of all except T. viride culture filtrates was significant on shoot biomass. There was a 9-56% reduction in shoot biomass due to foliar spray of culture filtrates on 1-wk old plants, while 2-wk old plants were comparatively tolerant to foliar spray treatments. In 2-wk old plants, foliar spray of only T. reesei culture filtrates significantly reduced the shoot biomass by 35% (Figure 1B). Root length was significantly reduced by 26% and 39% due to foliar spray of T. pseudokoningii and T. reesei, respectively, in 1-wk old A. fatua plants. Root biomass of A. fatua plants was also susceptible to foliar spray of culture filtrates at 1-wk growth stage. At this growth stage, foliar spray with culture filtrates of T. pseudokoningii, T. reesei and T. viride significantly reduced root biomass. None of the foliar spray treatments showed a significant effect on root length and biomass in 2-wk old plants (Figures 1C and D). The effect of foliar spray of culture filtrates of all the four Trichoderma spp. on various parameters of root and shoot growth of wheat was insignificant (Figure 2). DISCUSSION Culture filtrates of the four Trichoderma spp. tested: T. harzianum, T. pseudokoningii, T. reesei and T. viride, exhibited herbicidal activity against A. fatua. These finding are in agreement with those of the few studies carried out earlier in this field (Hutchinson, 1999; Heraux et al., 2005). The earlier studies regarding the herbicidal activity of Trichoderma spp. were restricted to the use of T. virens in combination with chicken manure (Hutchinson, 1999; Heraux et al., 2005). However, the present study reveals that the metabolites of other Trichoderma species also exhibit herbicidal activity. Trichoderma spp. produces a range of secondary toxic metabolites, including gliovirin, gliotoxin, viridian, and viridiol, of which viridiol is strongly phytotoxic (Jones and Hancock, 1987; Jones et al., 1988). Metabolites of T. harzianum exhibited herbicidal activity against A. fatua in both laboratory and foliar spray bioassays. Production of toxins belonging to the trichothecene class has been reported from T. harzianum (Sivasithamparam and Ghisalberti, 1998). Trichothecenes have been proven to be phytotoxic (Harris et al., 1999).These are sesquiterpenoid epoxides and represent a large family of secondary toxic metabolites produced by a variety of filamentous fungi (Ueno, 1980). Many of the toxic properties of the trichothecenes are attributed to their ability to inhibit protein and DNA synthesis (McLaughlin et al., 1977) and to induce apoptosis in eukaryotic cells (Okumwai et al., 1999). Recently, Gallo et al. (2004) isolated and characterized a putative trichodiene synthase (tri5) gene in T. harzianum required for trichothecene biosynthesis. Another compound trichosetin, a tetramic acid-containing metabolite produced in the culture of T. harzianum, is known to exhibit phytotoxic activity (Marfori et al., 2003). Vital staining of trichosetin-treated Nicotiana tabacum BY-2 cells, with rhodamine 123, showed a weaker green fluorescence compared to controls, indicating damaging effects on mitochondria. FDA-PI staining, to determine cell viability, indicated that cells of the trichosetin-treated roots were mostly dead (Marfori et al., 2003). T. harzianum also synthesises 6-pentyl-α-pyrone (6PP), a lactone with coconut-like aroma characteristics, is used in food and pharmaceutical products (Rocha-Valadez et al., 2006). This compound is also known to exhibit phytotoxic activity (Rocha-Valadez et al., 2006), and may derive from 6PP’s ability to bind to or permeate cell membranes (Poole and Whitaker, 1997). Culture filtrates of T. pseudokoningii proved highly toxic, resulting in significant growth suppression of A. fatua plants in both the laboratory bioassay and the foliar spray trial. T. pseudokoningii culture can produce complete cellulase enzymes, namely endoglucanase, cellobiohydrolase, and β-glucosidase (Ma et al., 1990),which may be responsible for damage to the cell wall,resulting in reduced plant growth. In addition, a low molecular weight peptide, termed short fiber generating factor (SFGF), derived from the culture supernatant of T. pseudokoningii, decomposes cellulose by oxidative reaction, which differs from the hydrolysis of cellulose by cellulases. The alcoholic hydroxyl groups in cellulose can be oxidized by SFGF, which leads to destruction of the hydrogen bond network in cellulose and cleavage of glycosidic linkages. Both effects led to the depolymerization of cellulose and the formation of short fibers, further reducing groups in residual cellulose, making the cellulose substrates more susceptible to hydrolysis by cellulases (Wang et al., 2003). Certain other bioactive chemical substances, such as acetophenone derivatives 2′,4′-dihydroxy-3′-methoxymethyl-5′methylacetophenone and 2′,4′-dihydroxy-3′,5′dimethylacetophenone (Astudillo et al., 2000),Pseudokonins KL III and KL VI (Rebuffat et al., 2000),and Harzianin HK VI (Rebuffat et al., 1996) have also been isolated from culture filtrates of T. pseudokoningii, and may be responsible for the herbicidal activity of the fungus. Culture filtrates of T. reesei were found to be comparatively less toxic than filtrates of T. harzianum and T. pseudokoningii in laboratory bioassays. Generally, only 100% culture filtrates of T. reesei exhibited significant effect against shoot and root growth of the target weed species and the effect of 50% culture filtrates were insignificant. However, in the foliar spray bioassay, the adverse effect of culture filtrates of T. reesei on 1-wk old A. fatua plants was similar to that of culture filtrates of T. harzianum and T. pseudokoningii. Trichodermin, a member of the trichothecene toxin family – a group of sesquiterpenes, is produced by T. reesei in culture medium. This secondary metabolite may be responsible for herbicidal activity against A. fatua. CONCLUSIONS The present study concludes that Trichoderma spp.namely T. harzianum, T. pseudokoningii and T. reesei produce certain secondary metabolites in the culture medium that exhibit herbicidal activity against A. fatua, a problematic weed of wheat. Further studies are in progress regarding the isolation and identification of potential herbicidal constituents in the culture filtrates of various tested Trichoderma species. LITERATURE CITED
Copyright 2011 - Chilean Journal of Agricultural Research The following images related to this document are available:Photo images[cj11024t2.jpg] [cj11024t1.jpg] [cj11024f2.jpg] [cj11024f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}