 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
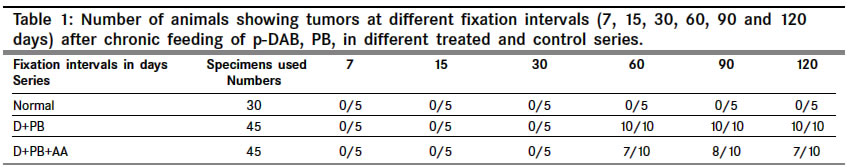
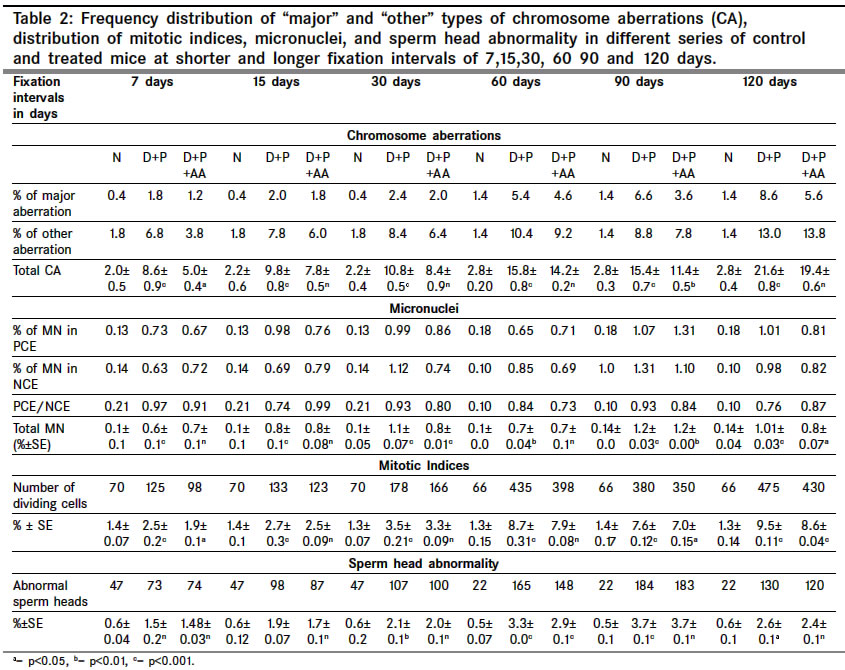
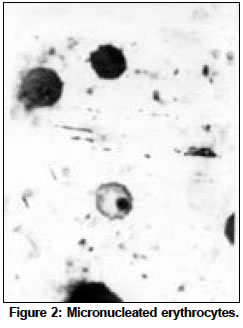
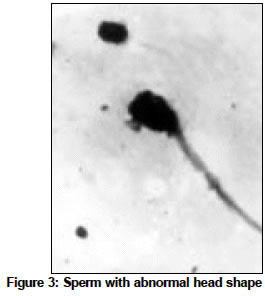
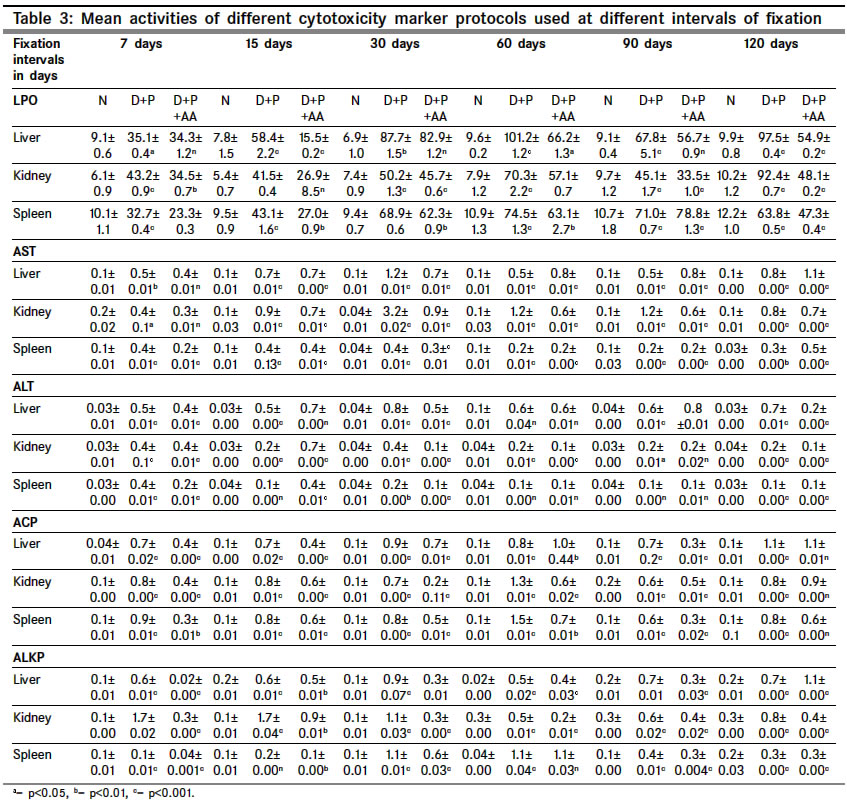
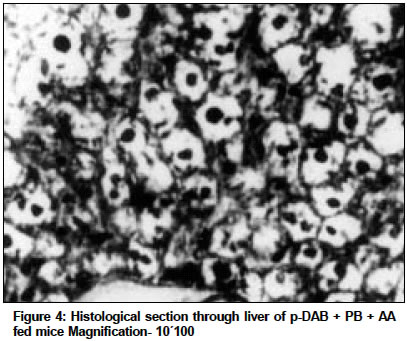
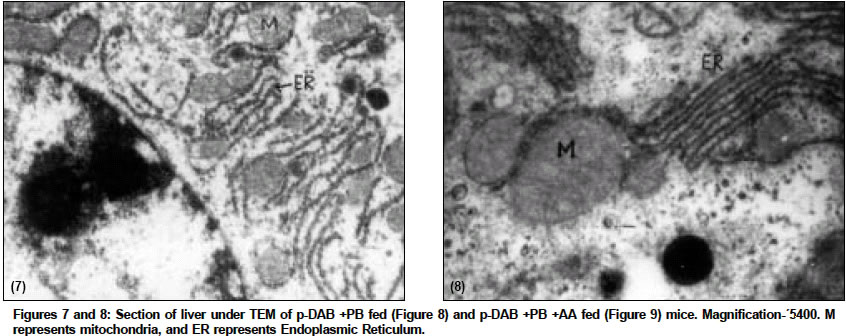
Indian Journal of Cancer, Vol. 41, No. 2, Apr-Jun, 2004, pp. 72-80 Original Article Protective action of an anti-oxidant (L-Ascorbic acid) against genotoxicity and cytotoxicity in mice during p-DAB-induced hepatocarcinogenesis Surjyo B, Anisur Rahman KB Cytogenetics Laboratory, Department of Zoology, University of Kalyani, Kalyani - 741235, West Bengal Correspondence Address:Cytogenetics Laboratory, Department of Zoology, University of Kalyani, Kalyani - 741235, West Bengal arkb@klyuniv.ernet.in; khudabukhsh_48@rediffmail.com Code Number: cn04013 ABSTRACT BACKGROUND: DNA damage from micronutrient deficiencies has been suggested as one major cause of cancer. Therefore studies involving vitamin supplementaion , particularly with those with anti-oxidant activity, in combating cancer have routinely been carried out in both in vivo and in vitro systems, but relatively much less in mice. AIMS: The present study examines if L-Ascorbic acid (AA; vitamin C) administration has any protective abilities in combating p-DAB induced hepatocarcinogenesis in mice at cytogenetical, biochemical, histological and ultra-structural levels. SETTINGS AND DESIGN: To test if AA had a protective action against genotoxicity, cytotoxicity and tissue damage in liver during p-dimethylaminoazobenezene (p-DAB) induced hepatocarcinogenesis in mice, a group of mice were chronically fed 0.06% p-DAB and 0.05% phenobarbital (PB) for a varying period of time (7, 15, 30, 60, 90 and 120 days). A sub-group of the p-DAB plus PB fed mice were also fed 1% L-ascorbic acid. Several assays were periodically conducted (at the six intervals of fixation) for determination of genotoxic (based on chromosomal, nuclear and sperm head anomalies), cytotoxic (based on the marker enzymes aspartate transaminase; AST, alanine aminotransferase; ALT; acid phosphatase; ACP; alkaline phosphatase; ALKP; lipid peroxidation; LPO); and tissue damaging (based on optical and electron microscopic studies of liver at day 60 only) effects in these different groups of mice as compared to normal healthy control. METHODS AND MATERIAL: Adult healthy mice of Swiss Albino strain, reared and maintained in the animal house of the Department of Zoology,. Kalyani University, under supervision of Animal Welfare Committee (which oversees ethical issues), served as materials for the present study. Widely practiced standard technique has been followed for each protocol. STATISTICAL ANALYSIS USED: The significance test between different series of data was conducted by student¢s t-test. RESULTS AND CONCLUSIONS: The results of all these studies indicated that AA had protective action against p-DAB induced hepatocarcinogenesis in mice. Keywords: Hepatocarcinogenesis, mice, p-dimethylaminoazobenzene; L-Ascorbic acid; anti-carcinogen, genotoxicity, cytotoxicity INTRODUCTION Many epidemiological and experimental studies indicate that diets rich in micronutrients may reduce the risk of cancer and mutation.[1],[2],[3],[4] Vitamins are very important as micronutrients in protecting macromolecules. Ascorbic acid (AA) is one such important micronutrient, which functions as a factor in several metabolic reactions.[4],[5] Further, DNA damage from micronutrient deficiencies has also been attributed as a major cause of cancer.[6] Therefore, various studies have been conducted during recent years to investigate the genotoxic and antigenotoxic properties of AA[7],[8],[9] in both in vitro and in vivo systems, though at a much lesser scale on in vivo system, particularly in mice.[10],[11],[12] However some conflicting role of ascorbic acid (AA) has been suggested in either inhibiting or enhancing carcinogenesis. While AA has been reported to have tumor enhancing effect in some mammalian models like guinea pig, rats[13],[14] and mice[15] its anti-carcinogenic role has also been advocated by others.[16],[17],[18],[19] The growth of solid form of Ehrlich ascites tumor has been reported to be significantly slower in mice maintained on distilled water supplemented with 1% AA.[20] Therefore we felt inclined to examine if this dose of 1% AA could also have a similar inhibitory effect in reducing p-DAB induced tumor induction/growth in liver and produce visible and quantifiable changes with regard to some cytogenetical and biochemical parameters. Further, visible changes or modulations brought forth by AA in mice, if any, were also carefully analyzed at histological and ultrastructural levels, which had not been done before in mice. METHODS An inbred strain of Swiss albino mice (Mus musculus), reared and maintained in the animal house of the Department of Zoology , under the supervision of the Animal Welfare Committee (which oversees ethical issues) , University of Kalyani, served as materials for the present study. Mice provided with food and water ad libitum served as normal controls. The food was made up of wheat, gram and powdered milk without any animal protein supplement. The chronic dietary method used by several workers[21],[22] in producing hepatic liver nodules and subsequent hepatocarcinoma was adopted. In the treated group, mice (45 samples) were allowed to take 0.06% p-dimethylaminoa-zobenzene (p-DAB) (Sigma, USA) mixed with food and 0.05% phenobarbital (PB) (Sigma, USA) instead of pure water. A group (45 samples) of p-DAB+PB fed mice were also administered orally with 0.06 ml of 1% L-ascorbic acid [SRL, India], i.e. 0.6 mg approx per mouse every day with the aid of a fine specially made pipette until they were sacrificed at day 7, 15, 30, 60, 90 or 120 of feeding. Slides for chromosomal assay were prepared by the conventional flame drying technique followed by Giemsa staining. Chromosome aberrations of various nature have been pooled into two categories: the "major" type comprising aberrations like break, fragments, ring, polyploidy, etc. and the "other" types comprising less significant aberrations like gaps, erosions, precocious centromeric separation, pycnosis, stretching, etc. A total of 500 bone marrow cells were observed, either 100 from each of 5 mice or 50 from each of 10 mice (longer intervals) of a set. For micronucleus (MN) preparation, clean grease free slides with smeared bone marrow cells were briefly fixed in methanol and subsequently stained with May-Grunwald (Sigma, USA) followed by Giemsa (Gurr, Germany). Approximately 5000 bone marrow cells, comprising both polychromatic erythrocytes (PCE) and normochromatic erythrocytes (NCE) were scored and the ratios between PCE and NCE ascertained. The mitotic index (MI) was assessed from the same slide that was scanned for MN. The non-dividing and dividing cells were recorded and their ratios calculated. For sperm head anomaly (SHA) study, the technique of Wyrobek[23] followed by Giemsa staining was adopted. The lipid peroxidation was estimated from the supernatant by the method of Buege and Aust.[24] 1 ml of sample (homogenate containing 0.1-0.2 mg of protein) was mixed thoroughly with 2 ml of TCA-TBA-HCL mixture. The absorbance of the sample was determined at 535 nm in a double beam spectrophotometer (Schimadzu, UV 180 model) against a suitable blank. The malonaldehyde concentration of the sample was then calculated. Similarly, the methods of Bergmeyer and Brent[25] was adopted for estimation of AST and ALT. For the study of acid and alkaline phosphatases, the method of Walter and Schutt[26] was followed. For preparation of histological slides of liver at day 60, the standard methodology using Bouin′s fixative and microtome sectioning has been followed. For electron microscopy of liver at day 60, the standard gold coating technique using critical point-drier (CPD-Biorad, Microscience Division, Warford England), sputter-coater (Agar Sputter Coater, Model 198, Stansted, United Kingdom) etc was adopted in case of scanning electron microscopy (LEO, 435VP, United Kingdom). For transmission electron microscopy, (TEM CM-10, Philips Microscope) the ultrathin sections (60-90 nm) were stained with uranyl acetate and lead citrate (Sigma, USA). The significance test between different series of data was conducted by student¢s t-test. RESULTS The number of animals showing tumor nodules in their liver on autopsy has been shown in [Table - 1]. Tumors started appearing at day 60 onwards, in all 10 out of 10 mice at each interval of fixation in the p-DAB+PB fed mice. However, in the p-DAB+PB+AA fed mice, 7 out of 10 mice at day 60, 8 out of 10 at day 90 and 7 out 10 mice at day 120 showed the formation of tumor nodules, indicating thereby positive protective effect of AA in combating tumorigenesis in mice. As compared to typical diploid metaphase spreads in normal control, there were various types of aberrations [Figure - 1] in the p-DAB+PB fed mice. The frequencies of chromosome aberration steadily increased in the p-DAB+PB fed series at all fixation intervals (p<0.001) except at day 90 where it was more or less same as at day 60 [Table - 2]. The frequencies of aberrations however were slightly suppressed in the p-DAB+PB+AA fed mice at all fixation intervals as compared to corresponding group of p-DAB+PB fed mice, though the differences were statistically significant only at day 7 (p<0.05) and day 90 (p<0.01). Micronuclei [Figure - 2] were visible in both polychromatic and normochromatic erythrocytes in the p-DAB+PB fed mice. The total frequencies of micronuclei in this series were also significantly higher than their corresponding controls at all fixation intervals (p<0.001, p<0.01 at day 60) [Table - 2]. The PCE/NCE ratio also indicated a more rapid rate of transformation of polychromatic to normochromatic erythrocytes in this series of mice at all fixation intervals. In the p-DAB+PB+AA fed mice the frequencies of micronuclei were marginally suppressed as compared to that of the p-DAB+PB fed mice at all fixation intervals except at day 7 and day 90 where the difference was insignificant. The frequencies of P/N ratio were also less in p-DAB+PB+AA fed mice as compared to that of p-DAB+PB fed mice at all fixation intervals except at day 15. The mitotic indices in p-DAB+PB fed mice gradually increased at successive fixation intervals till day 60 after which there was marginal decline at day 90 and a further increase at day 120, as compared to corresponding controls (p<0.001, [Table - 2]. The rate of mitotic division was strikingly inhibited in p-DAB+PB+AA fed mice as compared to p-DAB+PB fed mice at all corresponding fixation intervals, of which the differences were statistically significant at day 7, 90 (p<0.05) and day 120 (p<0.001). As compared to the sperm with normal head shape found in normal controls, various abnormalities in the head shape [Figure - 3] were encountered in the p-DAB+PB fed mice. The frequencies of sperm with abnormal head morphology increased successively in this series at all fixation intervals till day 90 after which it fell down to some extent at day 120, as compared to normal controls. The differences were statistically significant at day 15, 30 (p<0.01), day 60, 90 (p<0.001) and day 120 (p<0.05). There was a marginal suppression in increase of frequencies of abnormal sperm in the p-DAB+PB+AA fed mice as compared to that of p-DAB+PB fed mice at all fixation intervals except at day 7 see [Table - 2]. The differences were statistically insignificant at all fixation intervals except at day 60 (p<0.01). The lipid peroxidation (LPO) in liver of p-DAB+PB fed mice was successively increased till day 60, then fell down at day 90 only to rise again at day 120 see [Table - 3], as compared to normal controls. The differences were statistically significant at various levels (p<0.05 through p< 0.001). The LPO was found to be enhanced also in kidney and spleen tissue at different fixation intervals as compared to their control counterparts, usually showing a trend of rise till day 60, then it declined at day 90 either to rise again (kidney) at day 120 or to further decline (spleen) (p<0.001 to p<0.01, see [Table - 3]. On the other hand, there was a clear reduction of LPO in the p-DAB+PB+ AA fed mice, as compared to p-DAB+PB fed mice, with regard to all three organs, the differences being statistically significant in most cases. The aspartate transaminase (AST) activities in different organs of p-DAB+PB fed mice were increased at all fixation intervals as compared to their counterparts in control animals and the differences were statistically significant at various levels (p<0.05 to p<0.001, see [Table - 3]. There was a clear inhibition of AST activities in p-DAB+PB+AA fed mice as compared to p-DAB+PB fed mice, the differences were at statistically significant levels in the majority of cases (p<0.05 to p<0.001) except for liver and kidney at day 7 and kidney and spleen at day 90. The alanine amino transferase (ALT) activities in different organs of p-DAB+PB fed mice were appreciably increased in different organs as compared to their normal controls (p<0.05 to p<0.001, except for liver at day 60 and spleen at day 60 and 90; see [Table - 3] and the differences against normal control counterpart were statistically significant at various levels. In the p-DAB+PB+AA fed mice there was a considerable inhibition of ALT activity in some tissues at certain fixation intervals as compared to their counterparts in p-DAB+PB fed mice. However in statistical terms some of these differences were highly significant (p<0.001) while others were not. The mean activities of ACP were found to be elevated at highly significant levels (p<0.001) in the p-DAB+PB fed mice for all the tissues at all fixation intervals as compared to their counterparts in normal controls [Table - 3]. However there was a clear trend of inhibition in their activities in the p-DAB+PB+AA fed mice, noticeable in all the tissues and at all fixation intervals except for liver at day 60 and kidney at day 120. The activity of ALKP in the p-DAB+PB fed mice showed a similar increasing trend in all the tissues and at all fixation intervals (p<0.001 to p<0.01), as compared to controls, except for liver at day 15 [Table - 3]. In the p-DAB+PB+AA fed mice there was a striking inhibition of ALKP activity (p<0.01) in all the tissues and at all fixation intervals, as compared to p-DAB+PB fed mice, except for liver (p<0.001) and spleen at day 120. As compared to normal histological structures observed in liver of normal control mice, a careful study of liver sections in the p-DAB+PB fed mice revealed some destructive changes which include: excessive fibrosis in hepatic parenchyma, rapidly proliferating small cells, presence of vacuolated cells, nucleoplasm either condensed or absent and more ramification of blood capillaries. Some of these destructive changes were clearly less drastic in nature in the p-DAB+PB+AA fed mice, e.g. blood capillaries were few in number, fibrosis was less evident, and number of vacuolated cells was less in number [Figure - 4]. As compared to liver sections under scanning electron microscopy in normal controls fibrosis was evident in the liver of p-DAB+PB fed mice [Figure - 5] and there was evidence of tissue necrosis causing appearance of holes; red blood cells were scattered amongst the parenchymal tissue, suggestive of break-down of blood liver barrier. Further, cells appeared to be unhealthy. On the other hand, in the p-DAB+PB+AA fed mice [Figure - 6], fibrosis appeared to decrease though cells still looked unhealthy, hepatic boundaries were distinct, Kupffer cells were fewer in number and looked less drastically affected as compared to normal control. Similarly, as compared to the liver sections of normal control mice under transmission electron microscopy, liver sections of the p-DAB+PB fed mice [Figure - 7] showed several characteristic changes in their sub-cellular organelles. These included: i) mitochondria and lipid droplets were numerous; ii) some mitochondria with deformed and irregular arrangement of cristae were present; iii) nucleus was round with diverse nucleoplasm; iv) smooth endoplasmic reticulums (ERs) were broken at many places; and v) Kupffer cells were numerous and were activated probably due to increased secretion of lymphokines. On the other hand, in mice fed with p-DAB+PB+AA [Figure - 8], the effect appeared to be less drastic, e.g., number of mitochondria was fewer with their relatively unaltered orientation of cristae; Kupffer cells were less activated and few glycogen granules were evident; ERs were with fairly intact boundaries. DISCUSSION The simultaneous feeding of the carcinogens p-DAB (initiator) and PB (promoter) generally produced liver tumors in all mice treated for 60 days and more, while the incidence of tumor formation was apparently less, i.e., in 8 out of 30 mice fed with p-DAB+ PB+AA for longer periods did not show tumors on autopsy. It is generally accepted that p-DAB is metabolically degraded to N-methyl-4-aminoazobenzene (MAB), and then to 4-aminoazobenzene (AAB), which can bind to various liver proteins. The metabolites can also covalently bind with DNA.[27],[28] This binding is believed to be a major clastogenic and carcinogenic factor.[29] Although the metabolic fate of PB is not yet well established, it is now known as an inducer of cytochrome P450 in a wide range of organisms from bacteria to humans.[30] Cytochrome P450, a multigenic superfamily of microsomal haemoproteins, plays a key role in the metabolism of a wide variety of xenobiotics. One of the main targets of PB is the P450 2B sub-family and it is also an inducer of P450 3A subfamily in rats.[31] In the present study, an increasing number of chromosome aberrations, micronucleated erythrocytes and increase in mitotic activity were some notable features observed with the lapse of time, right from day 7 through day 120 of p-DAB + PB treatment. The chromosomal damage could have been produced as a direct action of the carcinogens and their metabolites on DNA by adduct formation, as suggested by Ohnishi[29] or else, some of them might actually have arisen as a result of indirect cytotoxic effects of the carcinogens. Micronuclei are generally formed of whole chromosome or of broken acentric chromosome fragments and result from a clastogenic as well as a spindle poisoning effect.[32] These are further enhanced by cell proliferation, which in turn is associated with tumor promotion and progression. Therefore, the carcinogens stimulate cell proliferation for their mitogenic action and more cytotoxic compounds are thereby produced, which in turn may cause regenerative cell growth.[33] An increasing number of sperm head abnormalities was also observed in the p-DAB+PB fed mice along with lapse of time. Mutagens that affect germ cells are also known to have spermatotoxic effects in mice[34] and generally cause an increase in the frequency of sperm head abnormality.[35] Evenson et al[36] demonstrated that such spermatotoxic effect might be due to alteration of testicular DNA and sperm chromatin structure. The feeding of AA could combat the spermatotoxic effects to some extent. As ascorbic acid has marked nucleophilic properties it might intercept the reactive metabolites thereby preventing their attack on nucleophilic sites on DNA, hence blocking adduct formation.[37],[38] Earlier, Levine[39] implicated the role of AA in the physiology of testis in regard to protein metabolism. Many enzymatic functions of (AA) are believed to be essential for the normal integrity and function of testis i.e. synthesis, development and maintenance of normal sperm.[40] Therefore, the protective role of AA on sperm head observed in the present study could also be due to its regulatory effect on protein metabolism and repair activities in the germinal cells. Quite appreciable changes also took place in their enzymatic activities. Some of these enzymes like ACP and ALKP have even been directly implicated to the extent of cellular damage and toxicity,[33],[41],[42],[43] as also in lipid peroxidation. Similarly, changes in AST and ALT have also been implicated to the altered transaminase activities necessitated in connection with combating/repairing protein damage and necrosis.[44] Several noticeable changes in histological patterns of liver were also noted. Similar destructive changes were also reported to occur in liver sections of guinea-pigs and rats studied through electron microscopy after these animals had been treated with carcinogens like nitrosamine and dimethylaminoazobenzene.[45] TEM also revealed significant destructive changes in sub-cellular organelles and ultra-structural organization of liver in the carcinogen fed mice, but signs of recovery were noticeable in the mice also fed AA. Along with the above genotoxic and cytotoxic changes in several tissue and histological changes observed in liver, visible external changes were also noticeable in form of tumor nodules in the liver of most mice treated with p-DAB + PB at day 60 and afterwards. Thus the results of the present study clearly demonstrated protective ability of AA against the tumorigenic and clastogenic potentials of p-DAB and PB in mice, and were in support of several other works that suggested the anti-cancerous effects of AA[16],[17],[18],[19] in other mammals. The mechanisms through which AA played a protective role could also be attributed to its free radical scavenging, anti-oxidant, apoptosis-inducing and nucleophilic properties as suggested by other workers.[19],[38],[44],[46] However, although the scavenging capacity of antioxidant vitamins has mainly been believed to prevent oxidative damage by neutralizing the free radicals,[12],[47] sometimes under a different condition these can also have co-genotoxic activity instead of normal anti-genotoxic action as reported in cultured mammalian cells[48] and in fish in vivo.[49],[50] ACKNOWLEDGEMENTS The authors are highly indebted to: Dr. T.C. Nag, Asst. Professor of Electron Microscope Facility at All India Institute of Medical Sciences, New Delhi, for kindly providing SEM and TEM facilities and reading the histo-pathological changes, and to the University of Kalyani, for financial support of the work.REFERENCES
Copyright 2004 - Indian Journal of Cancer The following images related to this document are available:Photo images[cn04013f7-8.jpg] [cn04013f4.jpg] [cn04013t3.jpg] [cn04013f1.jpg] [cn04013f2.jpg] [cn04013t2.jpg] [cn04013f3.jpg] [cn04013f5-6.jpg] [cn04013t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}