 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
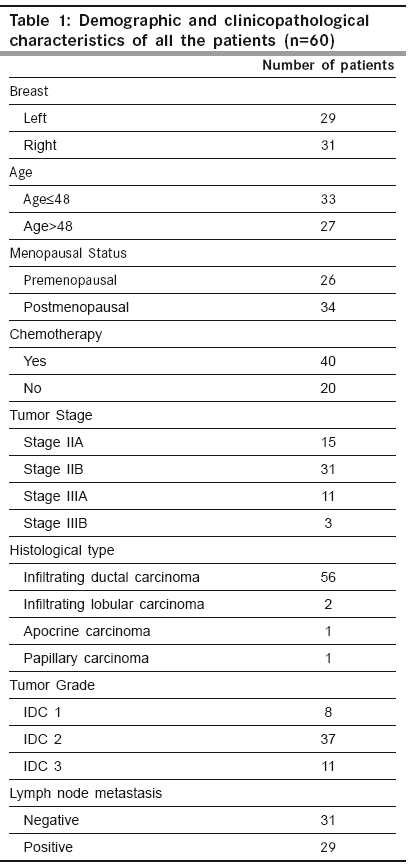
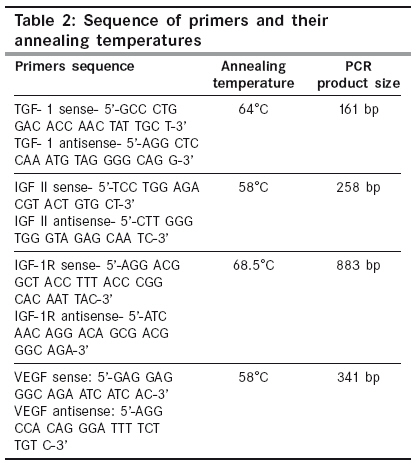
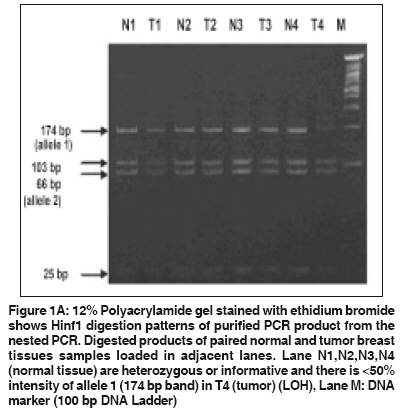
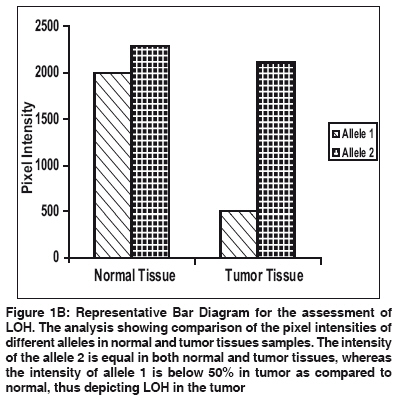
Indian Journal of Cancer, Vol. 46, No. 4, October-December, 2009, pp. 303-310 Original Article High frequency of loss of allelic integrity at Wilms' tumor suppressor gene-1 locus in advanced breast tumors associated with aggressiveness of the tumor Gupta S, Joshi K, Wig JD, Arora SK Department of Immunopathology, Postgraduate Institute of Medical Education & Research, Chandigarh Code Number: cn09072 PMID: 19749460 Abstract Background: The product of Wilms' tumor suppressor gene (WT1), a nuclear transcription factor, regulates the expression of the insulin-like growth factor (IGF) and transforming growth factor (TGF) systems, both of which are implicated in breast tumorigenesis and are known to facilitate angiogenesis. In the present study, WT1 allelic integrity was examined by Loss of Heterozygosity (LOH) studies in infiltrating breast carcinoma (n=60), ductal carcinoma in situ (DCIS) (n=10) and benign breast disease (n=5) patients, to determine its possible association with tumor progression.Methods: LOH at the WT1 locus (11p13) as determined by PCR-RFLP for Hinf1 restriction site and was subsequently examined for its association with intratumoral expression of various growth factors i.e. TGF-β1, IGF-II, IGF-1R and angiogenesis (VEGF and Intratumoral micro-vessel density) in breast carcinoma. Results: Six of 22 (27.2%) genetically heterozygous of infiltrating breast carcinoma and 1 of 4 DCIS cases showed loss of one allele at WT1 locus. Histologically, the tumors with LOH at WT1 were Intraductal carcinoma (IDC) and were of grade II and III. There was no correlation in the appearance of LOH at WT1 locus with age, tumor stage, menopausal status, chemotherapy status and lymph node metastasis. The expression of factor IGF-II and its receptor, IGF-1R was significantly higher in carcinoma having LOH at WT1 locus. A positive correlation was observed between the TGF-β1, VEGF expression and IMD scores in infiltrating carcinoma. Conclusions: The current study indicates that the high frequency of loss of allelic integrity at Wilms' tumor suppressor gene-1 locus in high-graded breast tumors is associated with aggressiveness of the tumor. Keywords: Ductal carcinoma in situ, insulin-like growth factor II, transforming growth factor β, vascular endothelial growth factor, Wilms′ tumor suppressor gene Introduction Normal growth and differentiation of the mammary gland depend on endocrine hormones that act in concert with locally produced growth factors such as insulin-like growth factors (IGFs) and members of transforming growth factor β (TGF-β) family. Multiple lines of evidence support the role of IGFs, acting through the IGF-1 receptor (IGF-1R), in both normal mammary growth and morphogenesis as well as in mammary tumor genesis. [1],[2] These genes are the targets of the Wilms′ tumor suppressor gene (WT1) product. In the normal kidney, WT1 suppresses the IGF-II and IGF-1R autocrine loop to modulate differentiation of renal epithelium. [3],[4] Loss of WT1 function contributes to Wilms′ tumor, via the activation of this loop, as demonstrated by the inhibitory effect of an antibody to IGF-1R on Wilms′ tumor in athymic mice, and in cell culture. [5],[6] Mutations and deletions of the Wilms′ tumor locus 11p13 have been found in sporadic Wilms′ tumor and are associated with congenital syndromes like Denysh-Drash syndrome, Beckwith-Wiedeman syndrome, WAGR syndrome. [7],[8] The studies have suggested that the loss of WT1 as a tumor suppressor gene may contribute to progression of other tumors like breast, kidneys etc. Based on the idea that, any mutation in the WT1 gene leading to LOH will result in over-expression of these growth factors and subsequent progression of breast carcinoma occurs by enhanced angiogenesis due to excessive production of VEGF. The present work is aimed to establish the frequency of LOH at the WT1 locus in DCIS and IDC in North Indian patients with the idea to determine the association of loss of allelic integrity with tumor progression. Material and Methods Acquisition of Samples: Sixty patients undergoing mastectomy were included in the study after informed consent. Of these, twenty six (43.33%) of the patients were premenopausal and the rest 34 (56.67%) postmenopausal. Forty patients (66.67%) were on neo-adjuvant chemotherapy, out of which 36 patients received post-operative adjuvant therapy. One patient had family history of breast carcinoma. The tissue samples were obtained from the surgical O.T. of the Nehru Hospital attached to Post Graduate Institute of Medical Education and Research, Chandigarh, India. Sixty biopsy samples of infiltrating breast carcinoma, 10 of ductal carcinoma in situ (DCIS) and 5 of fibroadenoma (benign breast disease) along with equal number of their adjacent normal tissues were included in the study. Tissue samples were snap frozen in optimum cutting temperature (OCT) compound and confirmation of diagnosis was made on a haematoxylin and eosin (H and E) stained frozen section. A histologically confirmed sample would typically include at least 80-90% tumor tissue in frozen sections. From one portion of tissues, DNA and total RNA were isolated and the other portion was processed for routine histopathological analysis. Relevant clinicopathological data including age, tumor stage, histological type of tumor, histological grade and axillary lymph node metastasis status was recorded in a predesigned format [Table - 1]. Tumor Staging was done according to TNM staging system laid down by the International Union against Cancer (UICC) and the American Joint Commission on Cancer staging. [9] However, tumor grade was assigned on HandE stained sections as described by Elston and Ellis. [10] Detection of LOH in tumors: Total DNA was extracted from the breast tissues by the method of Palmiter et al . [11] with some modifications. Extracted DNA was used as template in the external PCR to amplify a 952 bp fragment of WT1 gene locus. The primers (WT11: 5′-GCCTGGAAGAGTTGGTCTCT-3′; WT12: 5′-ACACAGTAATTTCAAGCAACGG-3′) were chosen from previously published sequences that flanked a polymorphic site on exon 9 of the WT1 gene. [12] The reaction mixture for PCR comprised of 0.8 µg of template DNA, PCR buffer supplied with the enzyme (consisting of 0.1 M Tris-HCl, pH 8.8, 15 mM MgCl 2, 0.5 M KCl and 1% Triton X-100), 250 mM dNTPs, 2.0 mM MgCl 2 , 0.5 pmole of each primer and 2 units of Taq DNA polymerase (Roche, Germany). After an initial denaturation at 95ºC for 5 min., the thermo-cycling was carried for 35 cycles with denaturation at 95 o C for 1 min., annealing at 60 o C for 1 min. and extension at 72 o C for 2½ min. in a thermocycler (Eppendorf, Germany). The last cycle was extended for 10 min at 72ºC. The PCR products were electrophoresed in a 1.25% agarose gel containing 0.5 mg/ml ethidium bromide and visualized on a UV-transilluminator (Fotodyne, USA). To increase the sensitivity of PCR, a nested PCR was performed using amplified product of external PCR as template and nested primers (R1: 5′-AATCAGAGAGCAAGGCATCGGG-3′; R2: 5′-CTTAAGAGCAGTGTGCCAGTG-3′) that amplified a segment of 225 bp. The primers were chosen from previously published sequences. [13] The amplified products were electrophoresed in 2% agarose gel containing 0.5 µg/ml ethidium bromide. Amplified PCR products of WT1 gene were purified using Invisorb® Spin PCRapid Kit (Invitek, Berlin). Twenty units of restriction enzyme Hinf1 (Roche, Germany) was used to digest the WT1 exon 9 at a polymorphic site in the purified PCR products from both external and nested PCR for 6 hrs. The digested PCR products were separated on 10% Polyacrylamide gel electrophoresis (PAGE) (in case of external PCR) and 12% PAGE (in case of nested PCR). Gel stained with ethidium bromide was visualized in a Gel Documentation System (Image Master VDS, Pharmacia Biotech). Heterozygous or informative profiles after restriction digestion of PCR products were identified by the presence of bands at 238 bp (allele 1), and 133/105 bp doublet (allele 2) in case of external PCR or 174 bp (allele 1) and 103/66 doublet (allele 2) in case of nested PCR, while the homozygous would contain either one of the alleles and would be non-informative profile. Band intensities of PCR product bands, as quantified by densitometric scanning, were obtained by taking the mean intensity value of pixels under a horizontal line drawn in the center of the bands representing each allele in the normal and tumor samples. LOH index was calculated by the formula:
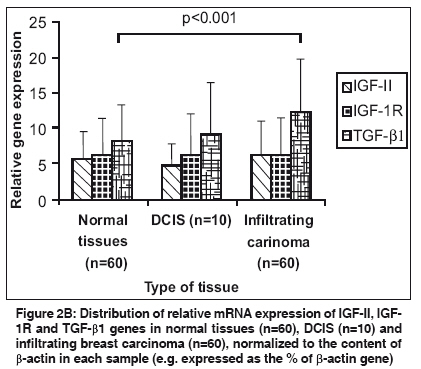
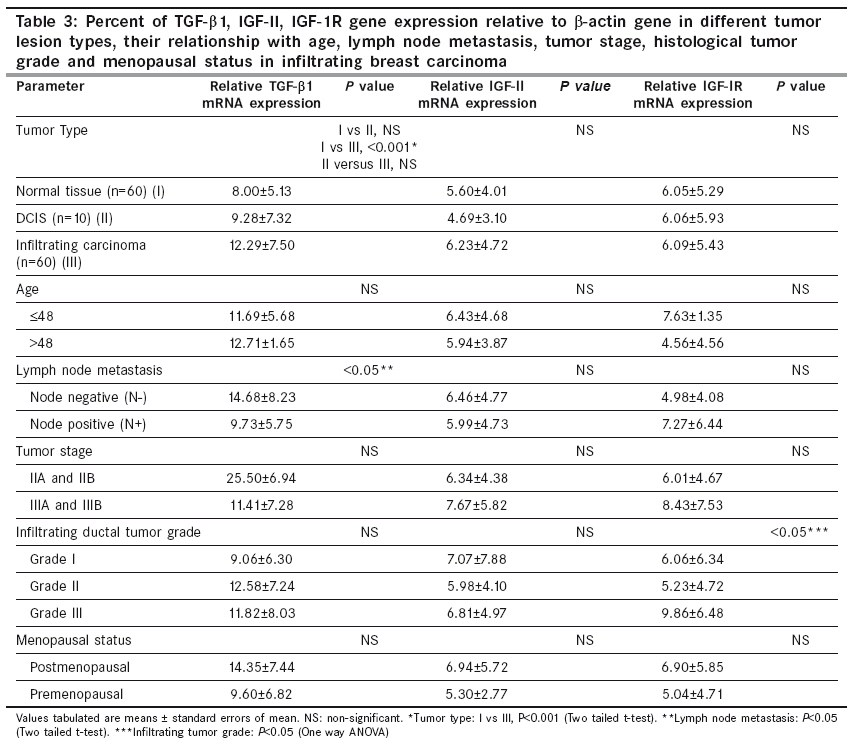
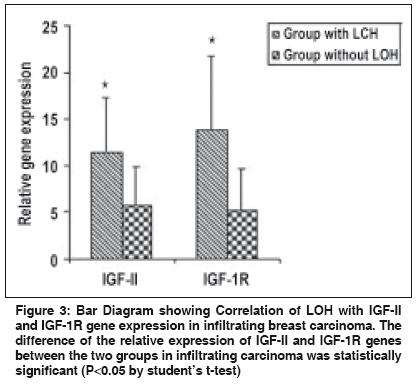
If LOH index was ≤0.6 or ≥1.7, it confirmed "Loss of Heterozygosity" (LOH) in tumor sample. Quantitative gene expression of IGF-II, IGF-1R, TGF-β1 and VEGF: The relative abundance of mRNA of each of these growth factor genes was semi-quantitatively estimated in each tissue by reverse-transcriptase PCR (RT-PCR). Total cellular RNA was isolated from the tumor tissue and their corresponding normal tissue using GITC method. [14] For RT-PCR, the complementary DNA strands (cDNA) were made from 2 µg of total RNA from each tissue using M-MLV reverse transcriptase enzyme (MBI fermentas, Lithuania) and random hexamers. For each PCR reaction, cDNA template was mixed to a standard reaction mixture consisting of 1 X reaction buffer, 1.5 mM MgCl 2 , 200 µM of each dNTPs, 0.5 pmoles of each oligonucleotide primer and 1.5 units of Taq DNA polymerase (Roche, Germany). Amplification of β-actin gene was used for normalization during quantitation as well as an internal quality control. The sequence of the primers, [15],[16],[17],[18] annealing temperatures and their product sizes are stated in [Table - 2]. Quantitation of TGF-β1, IGF-II, IGF-1R, and VEGF gene expression was done by comparing the signal intensities of RT-PCR product of these genes to those of β-actin gene from the same RNA sample using agarose gel electrophoresis containing ethidium bromide. The quantitative expression of each gene is given as the percent of constitutively expressed β-actin gene for each tissue sample. Immunohistochemistry for IMD: For microvessel density, the tissue sections were stained for endothelial cells using an anti-CD34 monoclonal antibody signifying neovascularization. Tissue sections (5 µ thick) were taken on poly-L-lysine coated slides. After deparaffinization and rehydration, the sections were subjected to microwave treatment in 0.01 M citrate buffer (pH 6.0) for 15 min., for antigen retrieval. The sections were stained overnight at 4°C with 10 -1 dilution of CD34 antibody (Dako, Denmark). A biotinylated anti-mouse antibody followed by streptavidin horseradish peroxidase was used sequentially for 30 min before color developement using chromogenic diamino benzidine (DAB 0.5 mg/ml) in the presence of 0.1% H 2 O 2 . Sections were then counterstained with haematoxylin. Sections of primary breast carcinomas in which primary antibody was omitted served as negative controls. Individual microvessels were counted in the area of highest vascularity. Any brown stained endothelial cell or cluster that was separate from other nearby microvessels was counted. [19] Results Allelic Loss at WT1 and clinicopathological significance: Evaluation of the Hinf1 polymorphism in the 60 infiltrating breast carcinomas revealed an overall rate of heterozygosity of 36.6% (22 of 60) with the presence of both the alleles. Of the 22 heterozygous tumors, 6 (27.2%) showed loss of either one of the alleles. In the DCIS group, of the 10 cases, 4 (40%) tumors were heterozygous, out of which one showed loss of one allele (allele 2). In the fibro adenoma group, all the 5 cases were homozygous. [Figure - 1a, 1b] illustrates the LOH-RFLP patterns of the nested PCR and bar diagram for the assessment of LOH. On analysis of LOH frequency in different histological types of invasive carcinoma, we found that all the six tumors with LOH were histologically classified as IDC and were of grade II and III. These findings suggest that accumulation of allelic losses at this chromosome locus might play an important role in tumor progression. There was however, no significant correlation of the LOH at WT1 locus with age, tumor stage, menopausal status, chemotherapy status and lymph node metastasis ( P >0.05). Intratumoral TGF-β1, IGF-II, IGF-1R expression and clinicopathological findings: The relative mRNA abundance of all these genes was found to be higher in tumor tissue as compared to normal tissue. Significantly higher levels of intratumoral TGF-β1 expression appear to be associated with the invasive phenotype of human breast carcinoma as compared to the non-invasive phenotype ( P < 0.001) [Figure - 2a, 2b]. [Table - 3] shows the correlation between various growth factors and each of the clinicopathological features. Analysis of variance by one way ANOVA revealed no significant difference except in IGF-1R levels ( P < 0.05), which were significantly higher in high grade tumors. There was inverse relationship of TGF-β1 gene expression with lymph node metastasis ( P < 0.05), which was higher (14.68±8.23) in the patients not having lymph node metastasis as compared to patients showing lymph node metastasis (9.73±5.75). There was no significant correlation between expression level of IGF-II and IGF-1R with any of the other clinicopathological features, including age, menopausal status or tumor stage. The relative abundance of VEGF mRNA in infiltrating carcinoma was (15.26±7.73) significantly higher than DCIS (8.31±5.60, P < 0.05) as well as normal tissue (5.40±3.22, p< 0.001). Similarly the IMD score was also significantly higher ( P < 0.001) in infiltrating tumors (145.18±71.33), than in DCIS [Table - 4]. Correlation of TGF-β1, IGF-II, IGF-1R with angiogenic factors in Infiltrating Breast Carcinoma: Correlation analysis was done to depict the association between expression of TGF-β1, IGF-II, IGF-1R, VEGF genes and IMD scores in infiltrating breast carcinoma tissues. A positive correlation was observed between the TGF-β1, VEGF expression and IMD scores. Correlation analysis of intratumoral expression of various genes viz. IGF-II vs IGF-1R, IGF-1R vs VEGF genes, VEGF vs IMD scores revealed a positive correlation, though it was not statistically significant. Correlation of LOH at WT1 with IGF-II, IGF-1R, TGF-β1 expression: Among infiltrating carcinoma group, IGF-II and IGF-1R expression was higher in the patient group which had LOH at the WT1 locus as compared to the group, which did not have LOH. The difference between the two groups was statistically significant ( P < 0.05) [Figure - 3]. In the DCIS group also, IGF-II and IGF-1R expression was higher in the patient, who had LOH at the WT1 locus. However, the difference in the TGF-β1 expression among the two groups was not statistically significant. Discussion Loss of tumor suppressor genes is an important mechanism in the development of cancer. It is widely accepted that progressive loss of tumor suppressor genes releases the cell from normal growth control, contributing ultimately to the malignant phenotype. In invasive breast cancer, LOH has been observed on many chromosomal arms, [20],[21],[22] making it difficult to ascertain which genetic events are crucial in oncogenesis. Most often, chromosome numbers 1, 13, and 17 show LOH. [23] In addition, chromosome 11 is frequently altered in human breast cancer. The loci 11p15 and 11q13 are reported to be putative tumor suppressor genes and have been linked with a poor prognosis in case of infiltrating breast carcinoma. [24] Domfeh et al .(2008) demonstrated WT1 expression in up to two-third of pure mucinous carcinomas and in approximately one-third of mixed mucinous carcinomas. But, no WT1 expression was seen in pure micropapillary carcinomas and mixed micropapillary carcinomas with ductal carcinomas, which indicated the putative role of WT1 as tumor suppressor gene. [25] In an endeavor to assess the frequency of LOH at WT1 locus in breast carcinoma specimen in patients from North India, we observed an overall heterozygosity rate of 36.6%. The frequency tended to be higher in infiltrating carcinoma (27.2%) as compared to carcinoma in situ (DCIS) and benign breast disease. All the six cases showing LOH at WT1 were histologically classified as infiltrating ductal carcinoma (IDC), whereas no allele loss was detected in any of the other histological types. Although the heterozygosity rate in our study with Indian population (36.6%) was lower than previously reported (59%) in Irish population by Fabre et al . [13] yet the frequency of LOH at WT1 locus in our study, was significantly higher (27.7%) than reported among Irish population (21.2%). Besides possibility of geographical differences, the high LOH frequency may reflect the superior separation of tumor cells from contaminating normal tissue in our study. Clinically advanced tumor stage and tumor grade are recognized prognostic factors in breast carcinoma. [26],[27] We found that all the tumors having LOH, were of high grade (grade II and III). Overall, these findings indicate the presence of putative tumor suppressor gene at chromosome 11 and also suggest that accumulation of allelic losses on this chromosome might play an important role in tumor progression. In contrast, there is no significant difference in LOH frequency with respect to age, tumor stage, menopausal status, chemotherapy or lymph node metastasis. Evidence for IGF action in the progression of WT has come from studies showing that inhibitory antibody directed against the human IGF and IGF-1R inhibits the growth of WT-derived cell lines in culture and in athymic mice. [5] IGF-1R protein and mRNA levels were elevated in Wilms′ tumor, and the WT1 protein also repressed the promoter of IGF-1R gene in vitro . [28] The growth factor genes like IGF-II, IGF-1R and TGF-β1 are known targets for transcriptional repression mediated by WT1 protein. The expression of IGF-II and IGF-1R was significantly higher in the group of patients who had LOH. A positive correlation in the expression levels indicates that both IGF-II and IGF-1R are co-regulated. This evidence suggests that the DNA binding domain of WT1 is inactivated in tumors and raises the possibility that this transcription-repression function is also inactivated. Moreover, the IGF-1R expression was found to be more in high grade lesions as compared to low grade lesions in our study suggesting that the overexpression of IGF-1R is associated with aggressive behavior of the breast tumor. Kucab and Dunn, [29] considered the potential role of IGF-1R in regulating breast cancer metastases by facilitating angiogenesis and lymphangiogenesis through the induction of VEGF. We also found a positive correlation between IGF-1R and VEGF expression in infiltrating breast carcinoma. These findings suggest that IGF-1R might have a potential role in promoting angiogenesis by the induction of VEGF expression in tumor cells. The TGF-β1 expression however, showed no correlation with LOH at WT1 in our study. A positive correlation was observed between the TGF-β1, VEGF expression and IMD scores. Correlation analysis of intratumoral expression of various other growth factor and their receptor genes viz. IGF-II vs IGF-1R, IGF-1R vs VEGF genes, VEGF vs IMD scores revealed a positive correlation, though it was not statistically significant. On the basis of this data, a unifying mechanism of the involvement of the IGF system in the progression of breast carcinoma can be formulated. During normal breast development, activation of WT1 gene expression represses synthesis of IGF-II and IGF-1R. The lack of WT1 function resulting from any of several mechanisms like LOH affecting the structure or expression of the WT1 gene, would leave the IGF-II autocrine loop intact leading to overexpression of this factor and possibly contribute to the inappropriate proliferation that leads to breast carcinoma. On the basis of findings in our study, we might speculate that the allelic loss at WT1 results in a truncated protein which fails to interact with its target DNA sequence. There remains possibility of other changes in WT1 gene besides LOH, which might also be responsible for similar effects. These results also suggest that the promoter of the IGF-II and IGF-1R gene constitutes a target for the inhibitory action of WT1. Paracrine activation of IGF-1R through binding of IGF-II results in an increased mitogenic action. This causes excessive proliferation of tumor cells which start expressing large amounts of TGF-β and VEGF, leading to increased vascularization and tumor growth. Although the direct effect of the WT1 gene product on the expression of IGF-II and IGF-1R gene has been shown on cell lines by in vitro studies, this is the first study showing indirect effect of the WT1 gene product in the surgically removed tumor tissue. These data, in no way by itself confirm that LOH at 11p is an obligate step in the development of all, or a specific subset of breast cancers, although the allelic loss at 11p13 may probably be one of the several steps in the genetic pathway of breast tumorigenesis. Larger studies would be helpful to resolve some of un-answered questions. References
Copyright 2009 - Indian Journal of Cancer The following images related to this document are available:Photo images[cn09072f2a.jpg] [cn09072t1.jpg] [cn09072f2b.jpg] [cn09072t2.jpg] [cn09072f1a.jpg] [cn09072f1b.jpg] [cn09072t4.jpg] [cn09072t3.jpg] [cn09072f3.jpg] |
| |||||||||


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}