 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
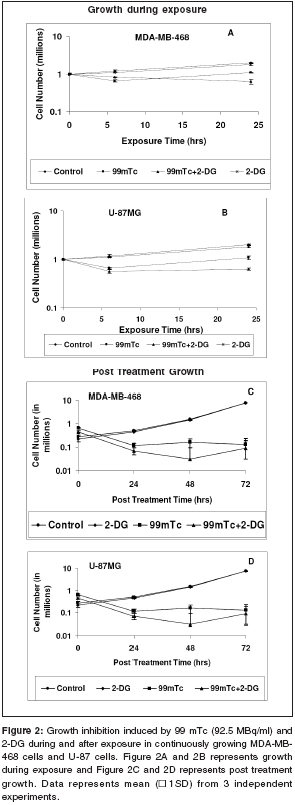
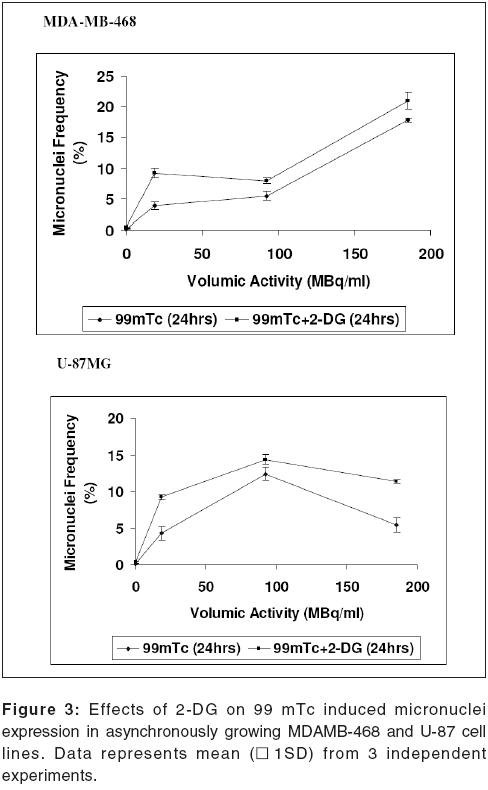
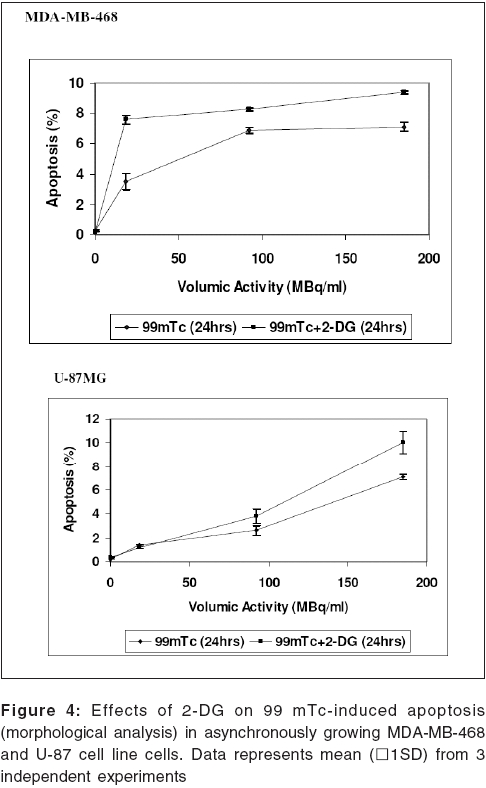
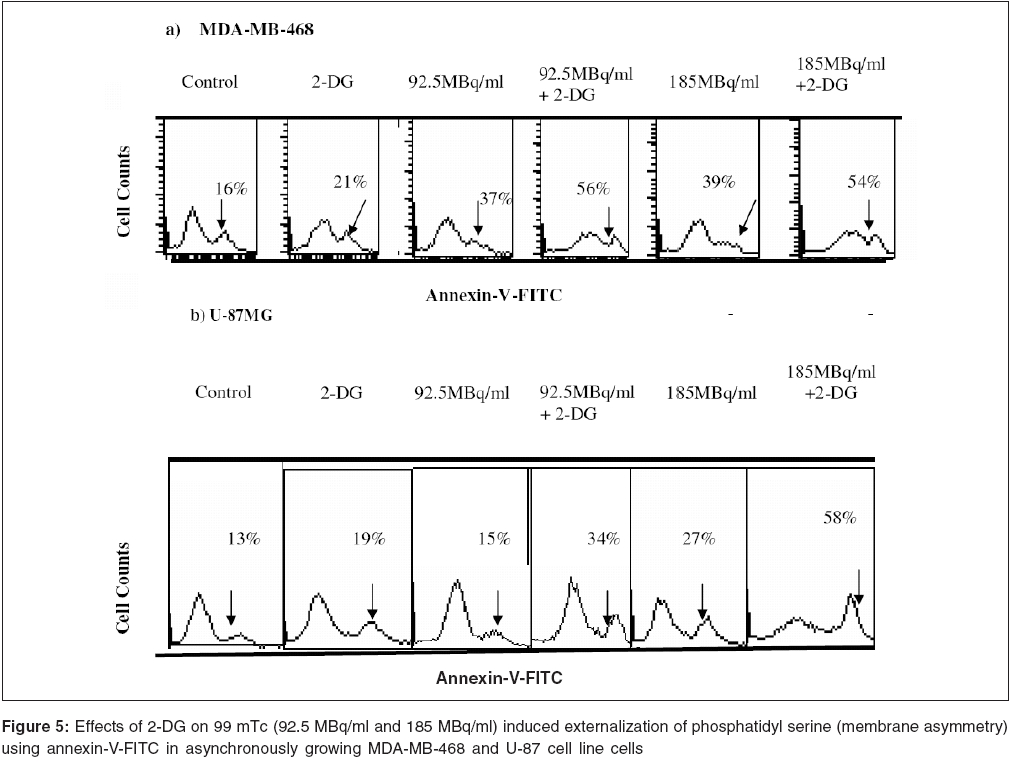
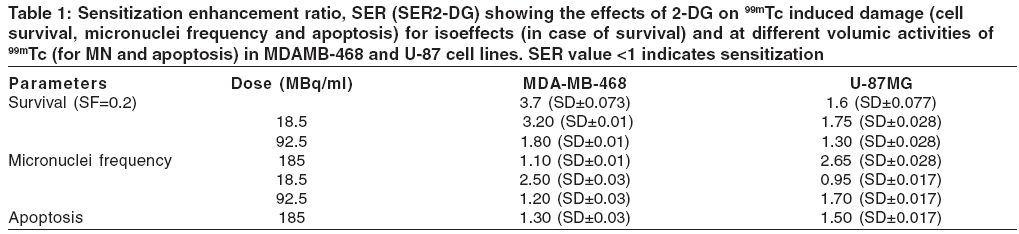
Journal of Cancer Research and Therapeutics, Vol. 2, No. 2, April-June, 2006, pp. 57-64 Original Article Enhancement of radionuclide induced cytotoxicity by 2-deoxy-D-glucose in human tumor cell lines Shrivastava V, Mishra AK, Dwarakanath BS, Ravindranath T Institute of Nuclear Medicine and Allied Sciences, Delhi Code Number: cr06013 Abstract The efficacy of targeted radiotherapy can be enhanced by selective delivery of radionuclide to the tumors and/or by differentially enhancing the manifestation of radiation damage in tumors. Our earlier studies have shown that the 2-deoxy-D-glucose (2-DG), an inhibitor of glucose transport and glycolytic ATP production, selectively enhances the cytotoxicity of external beam radiation in tumor cells. Therefore, it is suggested that 2-DG may also enhance the cytotoxic effects of radionuclides selectively in tumor cells, thereby improving the efficacy of radionuclide therapy. In vitro studies on breast carcinoma (MDA-MB-468) and glioma (U-87) cell lines, has been carried out to verify this proposition. Clonogenicity (macrocolony assay), cell proliferation, cytogenetic damage (micronuclei formation) and apoptosis were investigated as parameters of radiation response. Mean inactivation dose D (dose required to reduce the survival from 1 to 0.37), was 48 MBq/ml and 96 MBq/ml for 99 mTc, treated MDA-MB-468 and U-87, respectively. The dose response of growth inhibition, induction of micronuclei formation and apoptosis observed under these conditions, were correlated well with the changes in cell survival. Presence of 2-DG (5 mM) during radionuclide exposure (24 hrs), reduced the survival by nearly 2 folds in MDA-MB-468 (from 48.5 MBq to18.5 MBq) and by 1.6 folds in U-87 cells (from 96 MBq to 66 Mbq). These results clearly show that the presence of 2-DG during radionuclide exposure, significantly enhances the cytotoxicity, by increasing mitotic as well as interphase death. Further studies to understand the mechanisms of radio-sensitization by 2-DG and preclinical studies using tumor-bearing animals, are required for optimizing the treatment schedule. Keywords: Radionuclide, micronuclei frequency, 2-deoxy-D-Glucose, survival. Introduction Targeted radiotherapy involves the use of radiolabeled monoclonal antibodies (mAb) or peptidesthat bind to tumor-associated antigens or receptors, uniquely, or are over-expressed in tumor cells or tumor vasculature.[1] This therapy has certain advantages over external beam therapy, as it can selectively deliver radiation doses to the target tissues. However, small yet undesirable delivery of radiation to the non-target tissues, resulting in few immediate and late side effects[2],[3],[4] and the inherent radio-resistance of tumors, limit the success of this therapeutic modality. Therefore, approaches that can improve the selective delivery of radionuclide to the tumors and/or differentially enhance the manifestation of radiation damage in tumors, would significantly enhance the efficacy of radionuclide therapy. Ionizing radiation induces a number of DNA and non-DNA damages,[5] causing profound alterations in the gene expression and cell proliferation.[6],[7] It is well established, that the cellular processes leading to the repair and fixation of radiation-induced DNA damage (and possibly non-DNA damage as well, require a continuous supply of metabolic energy in the form of ATP[8],[9],[10],[11] produced by the respiratory and/or the glycolytic pathways.[4],[12],[11] Since tumor cells manifest a higher glucose uptake and derive a large part of their energy from the glycolytic pathway, it was postulated that inhibitors of glucose transport and glycolysis could differentially enhance radiation damage in tumors.[9],[4] Subsequently, it was demonstrated that 2-DG, a glucose antimetabolite, selectively inhibits DNA repair in cells with high rates of glycolysis (e. g. cancer cells), thereby enhancing the radiation damage in tumor cells in vitro and in animal tumors in vivo,[13],[14],[16],[17],[9],[18] following irradiation with low LET, as well as high LET external beam irradiation. On the other hand, a significant decrease in the radiation damage of normal cells has been observed under these conditions, possibly due to reduced damage fixation.[19],[20] The basic nature of DNA (and non-DNA] lesions induced by radionuclides and the molecular responses elicited by these lesions, show a great deal of similarity with external beam irradiation, although the rate of damage induction as well as kinetics of repair/fixation could vary, leading to qualitative as well as quantitative differences in the cellular radio responses. Therefore, it has been proposed that metabolic inhibitors such as 2-DG, could also modify the cellular responses to radionuclide- induced damage, by modulating the repair as well as damage-dependent cell proliferation and cell death processes. To verify this proposition, we have initiated studies to investigate the effects of 2-DG on the cytotoxicity induced by free as well as by radionuclides, conjugated to various specific vectors, that provide different types of radiation (γ and β emitters). In the present communication, wepresent our results on the free 99 mTc (gamma and auger emitter) treated human tumor cell lines.Materials and Methods Cell culture Cell survival: 99mTc and 2-DG treated cells were washed twice with HBSS, trypsinized and 100 to 1000 cells were plated in 60-mm petridishes and incubated at 37°C, in 5% CO2 humidified atmosphere (depending on the treatment), for 8-10 days. Colonies were fixed in methanol and stained with 1% Crystal violet. Colonies containing more than 50 cells were counted. To compare the extent of 2-DG induced sensitization or modification among different cell lines, sensitization enhancement ratio- SER, was calculated using formula SERSF 2-DG = D1/D2, where D1 and D2 are the required doses of 99mTc, without and with 2-DG, to obtain a surviving fraction of 0.2. (A value of SERSF 2-DG > 1 means an increase in the damage (sensitivity), while rSF 2-DG < 1 means a decrease). Cell growth Cytogenetic damage Air dried cells were stained with Hoechst-33258 at a final concentration of 10 mg/ml, in a buffer containing 50 mM sodium phosphate buffer, 10 mM Citric acid and 0.5% Tween-20 detergent. The binucleated cells with micronuclei, were scored with the help of a fluorescence microscope, using UV excitation. A minimum of 500 cells each from duplicate slides, were scored per group. SER (SER2-DG) was calculated using formula: SERMN Apoptosis Morphological analysis SERAPOP Phosphatidylserine externalization The percentages of annexin-V +ve and -ve cells, were estimated by applying appropriate gates. Statistical analysis Results
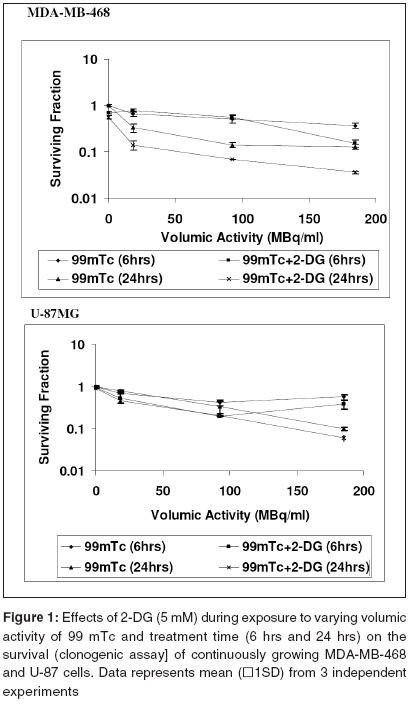
Cell survival Presence of 2-DG (5 mM) during radionuclide exposure (24 hrs), reduced the survival by nearly 2 folds in MDA-MB-468 (from 48.5 MBq to 18.5 MBq) and by 1.6 folds in U-87 cells (from 96 MBq to 66 Mbq). Under these conditions, 2-DG alone was minimally cytotoxic to the U-87 cells, while a 30% cell kill was observed in MDA-MB-468 cells for a 24 hrs exposure, suggesting that the cell kill observed with the combined treatments were clearly supra-addiditive, implying sensitization to 99mTc. However, presence of 2-DG during a 6hrs exposure to 99mTc, enhanced the cytotoxicity by 2.4 fold only at the higher dose, in MDA-MB-468 cells ( P _0.03), while 1.8 folds enhancement at 92.5 and 185MBq doses, was observed in U-87 cells ( P _0.0026 and 0.03 respectively). Cell proliferation Micronuclei Induction Apoptosis Presence of 2-DG during exposure to radionuclide, showed dose-dependent increase in the frequency of apoptotic cells, with maximum increase of nearly three folds at 18.5 MBq/ml in MDAMB-468 cells. On the other hand, in U-87 cells, induction of apoptosis was significantly higher (40%), at higher dose 185 MBq/ml [Figure - 4]. Under the present experimental conditions, 2-DG alone, did not induce apoptosis in both the cell lines [Figure - 4]. Phosphatidylserine externalization Discussion In the present studies, cellular radiation responses to damage caused by a radionuclide 99mTc (gamma rays and auger electrons), as well as modification of these responses by 2-DG, have been investigated in two human tumor cell lines. Results clearly show that the presence of 2-DG, an inhibitor of glycolytic ATP production,[18],[22],[27] during exposure to 99mTc, significantly enhances the cytotoxic effects of the radionuclide, by increasing the radionuclide- induced cytogenetic damage and apoptosis [Figure - 3],[Figure - 4]. Nearly 1.5 folds (U-87) and three folds (MDA-MB-468) increase in cell death, observed in these cell lines due to enhanced cytogenetic damage (micronuclei expression) (SER value up to 3.2), linked mitotic death and interphase death (apoptosis) (SER value up to 2.50). [Table - 1] clearly shows that 2-DG can enhance the cytotoxic effects of radionuclide (99mTc). However, quantitative differences were evident in the extent of sensitization [Table - 1] between the two cell lines, which could arise due to differences in the treatment induced, modification of different cell death pathways regulated by multiple gene products including p53, as well as differences in the degree of glycolysis between the two cell lines. Indeed, a positive correlation between the endogenous level of glycolysis and 2-DG-induced radiosensitization has been demonstrated, besides significant differences observed in the degree of glycolysis among different tumor cell lines.[15] Although a correlation between p53 status and radiosensitization by 2-DG has not been found so far,[15] it is pertinent to note that U-87 has a wild- type p53 status,[22] while the MDA-MB-468 carried a mutated p53 gene.[21] Induction of fewer lesions at low radiation doses, often result in insignificant changes in cell survival (shoulder region), whereas saturation of repair processes at higher doses leads to enhanced cell death. Quantitative differences observed in the extent of sensitization by 2-DG to 99mTc- induced cell death, at different doses in MDA-MB-cells, could partly arise on account of these reasons and which may differ from cell to cell. These observations are similar to our earlier results, on the radio-modifying effects of 2-DG, following external beam irradiation with 60Co gamma rays, in several human and murine tumor cell lines,[16],[17],[18],[22] where inhibition of DNA repair and cellular recovery processes have been demonstrated.[4],[17],[22],[28],[29] It is well known that as the dose rate of irradiation decreases, cell survival increases, which is predominantly due to the repair of radiation damage in the moderate dose rate range (from 200 cGy/min to 1cGy/min).[30] 99mTc is a gamma ray and auger electron emitting radionuclide and is homogeneously distributed in various sub cellular sites with 48% in nucleus, 17% in microsomes, 20% in mitochondria and 15% in the membrane, so that it can deliver continuously decreasing dose rates as a function of time, due to disintegration of the radionuclide, uniformly throughout the cells.[31] Therefore, varying the volumic activity as well as exposure time, actually represents cumulative dose effects of continuously decreasing dose rate. Nearly 60% cell kill was observed with a 24 hexposure at 18.5 MBq/ml of the radionuclide in MDA-MB-468 cells [Figure - 1] and the value of the surviving fraction (0.33) was similar to the SF observed at 5 Gy, at a dose rate of 0.076 Gy/min, delivered from an 60 Co source of external beam irradiation [data not shown] . At this dose rate, repair during irradiation is the predominant cause of the enhanced survival, as compared to high dose rate (2 Gy/min) irradiation.[30] Since it is reasonable to expect that the repair of DNA damage occurs during exposure of cells to radionuclide [24h], inhibition of repair and/or recovery processes, is expected to enhance cell death. Indeed, inhibition of DNA repair and cellular recovery processes by 2-DG, following damage caused by external beam irradiation, has been reported.[17],[22],[28],[29],[32] Under similar conditions, 2-DG alone had minimal cytotoxicity in U-87 cells, whereas a growth-inhibitory effect resulting in 30-40% reduction in cell number, was observed in MDA-MB-468 for 24 hr exposure, similar to the earlier results reported in this cell line.[23] Variation in the 2-DG- induced cytotoxicity has been previously reported and is related to the complexicity of uptake in irradiated cells and to the levels of pro-apoptotic or anti apoptotic proteins.[33] However, alterations in other post-irradiation responses viz. cell cycle perturbation and apoptosis, that influence survival, also contributes to the cytotoxicity. An alternative explanation for the enhanced radionuclide- induced cell death by 2-DG, could also be the redistribution of the radionuclide in the intracellular milieu, due to altered metabolic state, induced by 2-DG, as has been shown for 64Cu-ATSM in EMT-6 rat breast tumors.[34] However the uptake and intracellular distribution of radionuclide used for current study (99mTc), is known to be neither energy- dependent nor carrier- mediated.[35] Mitotic (cytogenetic damage) and interphase death (apoptosis), are two predominant modes of cell Death, that contribute to the loss of survival. Mitotic death is linked to the cytogenetic damage, expressed as chromosomal aberrations in the metaphase and manifest in the form of micronuclei-formation in the post mitotic daughter cells, which arise from the residual DNA damage, following induction and repair of DNA lesions. On the other hand, apoptosis is induced by both, membrane as well as DNA damage, which are observed as early as few hours in certain cell systems[36],[37] and after many hours or days in epithelial tumor cells.[24] It is also suggested that mitotically dead cells undergo apoptosis as a secondary response, often referred to as delayed apoptosis and observed 1-2 days after irradiation.[7],[38],[39] The surviving fraction of 0.34 and 0.1 observed following exposure to 95.2MBq/ml and 182.5 MBq/ml of 99mTc in U-87 cells, is comparable to an absorbed dose of 2Gy and 5Gy of external beam gamma rays, at 0.076Gy/min dose rate, where mitotic death linked to cytogenetic damage in the form of micronuclei expression (20-25%), is the primary cause of cell death, with minimal (< 5%) delayed secondary apoptosis.[7] Interestinglyhowever, the dose (activity) dependent changes in the micronuclei expression in U-87 cells, showed a decrease at 182.5MBq/ml, as compared to 92.5MBq/ml [Figure - 4]. This was accompanied by a concomitant increase in the fraction of apoptotic cells, implying a possible quantitative difference in the mitotic vs. apoptotic death. The lack of correlation between micronuclei frequency and apoptosis to the loss of clonogenecity, suggests the contribution of other modes of cell death like necrosis and non apoptotic delayed lethal mutations, as suggested by Little et al.[40],[41] However, significant differences could not be observed in the modification of these two responses viz. apoptosis and mitotic death by 2-DG, suggesting that the primary effects of 2-DG could be the modification of the residual DNA (and possibly non DNA) damage, on account of inhibition of repair processes. Indeed both DNA and non DNA damages can induce apoptosis, that depend on several gene products, which facilitate the progression of apoptosis through p53 dependent and independent mechanisms,[42] involving caspase- dependent and independent pathways. Therefore, the possibilities of 2-DG- induced alterations in the signalling mechanisms, could also be partly responsible for the enhanced apoptosis observed. Induction and expression of several genes related to various signaling and metabolic pathways have been recently reported, following the exposure of cells to radiation and 2-DG.[43] In RIT, the local tumor control achieved, is due to contributions from the radionuclide and/or toxicity compound associated with the vectors. Therefore, radio-modifiers which are able to enhance the manifestations of radionuclide- induced damages, can further enhance the efficacy of radionuclide therapy. Results of the present studies clearly showed that presence of 2-DG during radionuclide exposure, significantly enhances the cellular damages and imply that 2-DG can enhance the efficacy of radionuclide therapy. Preclinical studies in tumor-bearing animals, besides investigations on the effects of the combined treatment (radionuclide + 2-DG) on the damage to normal cells and tissues, are required before contemplating clinical studies. References
Copyright 2006 - Journal of Cancer Research and Therapeutics The following images related to this document are available:Photo images[cr06013f3.jpg] [cr06013f5.jpg] [cr06013f1.jpg] [cr06013f4.jpg] [cr06013f2.jpg] [cr06013t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}