 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
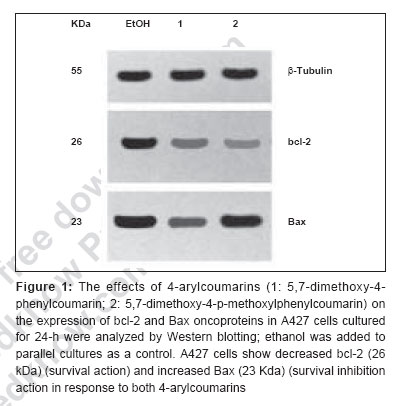
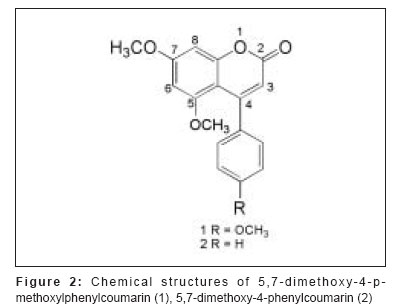
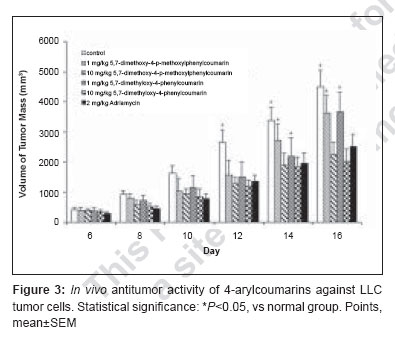
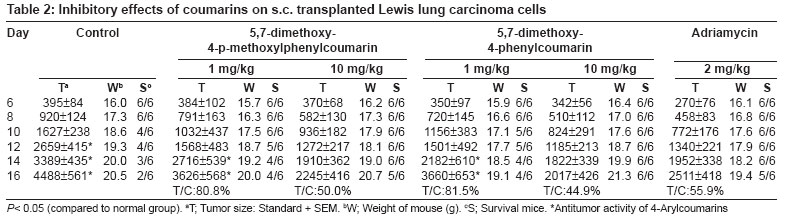
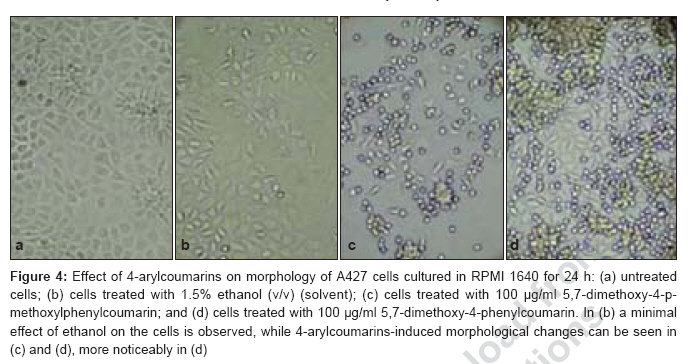
Journal of Cancer Research and Therapeutics, Vol. 3, No. 2, April-June, 2007, pp. 86-91 Original Article Antitumor activity of 4-Arylcoumarins from endophytic Streptomyces aureofaciens CMUAc130 Thongchai Taechowisan, Chunhua Lu, Yuemao Shen, Saisamorn Lumyong Department of Microbiology, Faculty of Science, Silpakorn University, Nakorn Pathom - 73000 Code Number: cr07022 Abstract In a search for antitumor agents, we carried out a screening of 4-arylcoumarins isolated from endophytic Streptomyces aureofaciens CMUAc130, by examining their possible inhibitory effect on the growth of s.c. transplanted Lewis lung carcinoma (LLC) in BDF-1 mice by intraperitoneal (i.p.) administration. The 4-arylcoumarins showed antitumor activity with T/C values of 80.8 and 50.0% at doses of 1 and 10 mg/kg of 5,7-dimethoxy-4-p-methoxylphenylcoumarin treatment, respectively and 81.5 and 44.9% at doses of 1 and 10 mg/kg of 5,7-dimethoxy-4-phenylcoumarin treatment, respectively, compared to adriamycin, which was used a positive control, with T/C value of 55.9% at 2 mg/kg. Furthermore, we investigated the possible effects of these compounds on expression of the bcl-2 and Bax oncoproteins in A427, a human lung cancer cell lines. The cells were cultured in vitro for 24h in RPMI 1640 with 1.5% (v/v) ethanol, 100 µg/ml 5,7-dimethoxy-4-p-methoxylphenylcoumarin or 5,7-dimethoxy-4-phenylcoumarin. Viability was determined by an MTT assay. Total protein was extracted from cell lysates and the bcl-2 and Bax oncoproteins were identified. Western blotting showed a decrease in bcl-2 and an increase in Bax in A427 cell cultured with 5,7-dimethoxy-4-p-methoxylphenylcoumarin or 5,7-dimethoxy-4-phenylcoumarin. We conclude that 5,7-dimethoxy-4-phenylcoumarin is a more potent inhibitor of cell proliferation than 5,7-dimethoxy-4-p-methoxylphenylcoumarin and has more marked effects on oncoprotein expression. Keywords: 4-arylcoumarins, antitumor activity, endophytic streptomyces, Lewis lung carcinoma Introduction Coumarins are synthesized by numerous green plant species [1] and by some bacteria and fungi and they have low toxicity in human. They exhibit antibacterial, [2] antioxidant, [3] anticancer [4] and antiallergy activities. [5] Recent report describe that 4-hydroxycoumarin derivatives have antiproliferative effects against five human tumor cell lines (A549 (nonsmall cell lung carcinoma), SK-OV-3 (ovarian carcinoma), SK-MEL-2 (melanoma), XF498 (CNS carcinoma) and HCT-15 (codon carcinoma)) [6] and act directly on malignant cells reducing tumor size [7] but the mechanisms involved have not been described in detail. In a preliminary communication, we have reported the isolation of 5,7-dimethoxy-4-p-methoxylphenylcoumarin and 5,7-dimethoxy-4-phenylcoumarin from the crude extraction of Streptomyces aureofaciens CMUAc130 culture. [8] In the course of searching for the antitumor agent from these compounds, we investigated the antitumor effect of these compounds on Lewis lung carcinoma (LLC) cells in vivo and the biochemical mechanisms of bcl-2 and BAX expression in human lung cancer cell lines, A427 (adenocarcinoma). The 26 kDa membrane protein bcl-2 may extend the life of the cell by blocking free oxygen radicals, inhibiting cytochrome c released from mitochondria and suppressing caspase-9 activation. [9] This would cause oncogenic mutations to accumulate in the normal cell, which may facilitate carcinogenesis. Over-expression of bcl-2 may also delay the response of tumor cells to applied therapies. [10],[11] Over expression of Bax, a 23 kDa membrane protein, results in the formation of homodimers and facilitates the production of channels in the outer mitochondrial membrane and loss of mitochondrial membrane potential that allows cytochrome c release. In the cytoplasm, cytochrome c binds Apaf-1 in the presence of ATP and activates inductor caspase-9, which in turn activates executor caspases that cleave death substrates, key cytoplasmic and nuclear proteins. [12],[13] Coumarins bind nonspecifically to cell surface structures like receptors. They function as first messengers, inducing changes in signal transduction pathways involving GTP-binding proteins. Therefore, coumarins are able to mediate modifications of downstream signals leading to growth (e.g. cell-growth inhibition) and possibly decreasing bcl-2 expression. It has been shown that other coumarin derivatives exert their antiproliferative effect by modulation of the cell cycle, [14] although the mechanism responsible for this activity has not yet been clarified. Accordingly, our findings indicate that 5,7-dimethoxy-4-p-methoxylphenylcoumarin and 5,7-dimethoxy-4-phenylcoumarin may display antitumor activity by inhibiting progression through the G0/G1 and S phases of the cell cycle. The Bcl-2 family regulates the fate of normal cells and comprises approximately eight members divided into two groups. Group 1 includes bcl-2 and bcl-XL, which has a survival action; group 2 includes Bax, Bad, Bid and Bak, which has a survival inhibition action. [9] The susceptibility of a cell to adverse stimuli is determined in most cases by the dimerization of Bcl-2 family proteins through the highly conserved BH1, BH2 and BH3 domains. [15],[16] The loss of survival inhibition Bax proteins or overexpression of the proteins that block them, results in resistance to treatment; this was observed by Pezzella et al. , [17] during a study of apoptosis inhibition in non-small cell lung carcinoma. It has also been reported that an increase in Bax expression sensitizes cells to chemotherapeutic agents. [18] The purpose of the present study was to determine whether 5,7-dimethoxy-4-p-methoxylphenylcoumarin and 5,7-dimethoxy-4-phenylcoumarin alter the tumor proliferation in vivo and the expression of the Bcl-2 oncoprotein family. Materials and Methods Organisms and compounds Structure elucidation of the compounds Tumor transplantation: Lewis lung carcinoma (LLC, mouse) cells were maintained as monolayer cultures in RPMI 1640 medium supplemented with 10% fetal bovine serum, sodium bicarbonate (2 g), pencillin G (100,000 units) and streptomycin (100 mg). A suspension of 2.5x10 5 cells/0.1 ml/mouse in 0.85% NaCl solution was inoculated subcutaneously into the left flank of female BDF-1 mice (four weeks of age, weighing 14 to 16 g) for the subcutaneous tumor assay. [22] The purified compounds, 5,7-dimethoxy-4-p-methoxylphenylcoumarin and 5,7-dimethoxy-4-phenylcoumarin were suspended in 0.85% NaCl solution (0.1 ml) and administered intraperitoneally (i.p) once daily for consecutive day 1 to day 14 to mice. Control animals were given 0.1 ml of 0.85% NaCl solution by i.p. injection. Each treatment consisted of three replicates. The experiment was repeated twice. Tumors were measured each alternate day using a vernier caliper from the initiation of treatment to the time when grosses ulceration of the tumor was developed in control mice. The tumor size was calculated as: Tumor vol. (mm 3 ) = 0.5xaxb 2 Where a is the longest diameter and b is the shortest diameter. [22] The effects by treatments were represented as follows: T/C (%) = (mean value of a treated group/mean value of a control group) x 100. The significance of differences between the experimental group was calculated by Dunnett′s t test. P < 0.05 was considered significant. Expression of bcl-2 and Bax in cancer cell line by purified compounds. Human lung carcinoma cell line used was A427 (adenocarcinoma) ATCC (American Type Culture Collection, MA, USA). For controls we used HeLa cells, which express the Bax oncoprotein and isolated human peripheral blood lymphocytes from healthy donors, which express the bcl-2 protein. These cells were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum and maintained at 37˚C in a humidified 5% CO2 atmosphere until 80-85% confluence was reached. They were harvested by using 0.25% Trypsin and their viability was determined by Trypan blue exclusion. A427 cells were seeded at a viable density of 1 x 10 7 in 10 ml RPMI 1640-supplemented medium. Untreated cell lines or cells in 1.5% (v/v) ethanol (solvent), 100 µg/ml 5,7-dimethoxy-4-p-methoxylphenylcoumarin or 100 µg/ml 5,7-dimethoxy-4-phenylcoumarin, were cultured for 24 h. Each culture was photographed at 200x with a Nikon H-III Power, FDX-35 inverted microscope to reveal ethanol and purified compounds-induced morphological changes. MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay %IC = [(1- cells with compound/cells in solvent)100] Protein isolation and Western blot analysis. Cells from each culture were lysed to extract total proteins by the TRIZOL method. [24] Brielfly, RNA and DNA were removed from the cell lysates and total proteins were precipitated from the final supernatant by adding 0.2 ml isopropanol per milliliter of TRIZOL used initailly. The precipitate was washed with 0.2% guanidine in 95% ethanol, then dissolved in 1% SDS and stored at -20˚C until it was analyzed. The total protein concentration was determined by the Petersen method. [25] Samples of total protein (30 µg/well) were separates on 10% SDS-PAGE, transferred to nylon membranes (Hybond-C Amersham Life Technologies) and incubated with the first antibody: anti-bcl-2(monoclonal antibody clone Bcl-2-100, labeling 26 kDa bands), anti-Bax (monoclonal antibody clone 2D2, labeling 23 kDa bands) (both from Zymed Laboratories, Inc. San Francisco, CA USA) or anti-β -tubulin (Monoclonal antibody clone Tub 2.1, labeling 55 kDa bands) (Sigma-Aldrich Inc, St. Louis MO, USA). The second (antimouse) antibody tagged with alkaline phosphatase was added with a 5-bromo-4-chloro-5-indolyl phosphate toluidine/nitroblue tetrazolium substrate as a color development reagent (Roche Diagnostics Co, Molecular Biochemical, Indianapolis, IN, USA). The band intensities were quantified by densitometry using RFLP scan software (Scanalytics, v. 2.1, 1994; Scanalytics, a division of CSPIMA, USA). The bcl-2 and Bax band values were normalized to the β -tubulin value (value = 1). Western blotting assays were repeated at least three times and a representative gel is included as [Figure - 1]. Data in [Table - 1] indicate the mean and standard deviation of three normalized densitometry values. Results and Discussion We have studied physiological roles of 5,7-dimethoxy-4-p-methoxylphenylcoumarin and 5,7-dimethoxy-4-phenylcoumarin for the development of biologically active substances. [8] Although there are many coumarin derivatives have special ability to scavenge reactive oxygen species-free radicals, such as hydroxyl radicals, superoxide radicals or hypochlorous acid and to influence processes involving free radical-injury. [26] They have also been found to inhibit lipid peroxidation and to possess vasorelaxant, [27] anticoagulant activities, [28] antiinflammatory/antioxidant activity. [29],[30],[31] Moreover they showed potent in vitro antitumor activity. [6] We selected 5,7-dimethoxy-4-p-methoxylphenylcoumarin and 5,7-dimethoxy-4-phenylcoumarin with a potent antitumor effect on LLC cells in vivo and the biochemical mechanisms of bcl-2 and BAX expression in A427. [Figure - 2] shows chemical structures of 5,7-dimethoxy-4-p-methoxylphenylcoumarin and 5,7-dimethoxy-4-phenylcoumarin. The purified compounds, 5,7-dimethoxy-4-p-methoxylphenylcoumarin and 5,7-dimethoxy-4-phenylcoumarin were evaluated for antitumor activity against subcutaneous transplanted LLC cells, in vivo . These compounds, when given intraperitoneally once daily for the 14 days at dosages of 1 and 10 mg/kg, inhibited tumor growth with T/C values of 80.8 and 50.0% for 5,7-dimethoxy-4-p-methoxylphenylcoumarin treatment, respectively and 81.5 and 44.9% for 5,7-dimethoxy-4-phenylcoumarin treatment, respectively, compared with that of the control [Figure - 3]. When the sample was intraperitoneally administrated once daily for 14 days, the body weights continued to until the fifth day after injection, following gradual increase and weighing almost same as body weights in control group [Table - 2]. Coumarins have been studied for their binding properties to tumor cell membranes in vitro , for their ability to inhibit cell proliferation in cells dependent on the over-expression of oncogenes. In order to measure the interaction, intact target cells were radioactively labeled with coumarin. Metastatic breast carcinoma cells showed high-affinity binding properties, where an adenocarcinoma raised from a human colorectal primary tumor exhibited low affinity to coumarin. The binding properties of coumarin to target cells are enigmatic, possibly due to biological variability. [32] Diffusion of coumarin molecules through the cell is determined by the phospholipid composition of the plasma membrane. Thus, the level of affinity of coumarins for membrane phospholipids constitutes the driving force that allows the entry of coumarins into the cell. [Figure - 4]a shows the untreated A427 cell line culture, while [Figure - 4]b shows this culture treated with ethanol. In both cases, the cells grew separately, adopting irregular forms with cytoplasmic elongations. [Figure - 4]c and d is the culture treated with 5,7-dimethoxy-4-p-methoxylphenylcoumarin and 5,7-dimethoxy-4-phenylcoumarin, respectively. Small, round cells tending to group together were observed in both these cultures and are particularly prevalent in [Figure - 4]d. No morphological changes were induced by ethanol. An MTT assay was performed to determine the inhibition of cell proliferation in the two cell lines cultured for 24-h with 100 µg/ml 5,7-dimethoxy-4-p-methoxylphenylcoumarin or 5,7-dimethoxy-4-phenylcoumarin or with 1.5% (v/v) ethanol (solvent control). [Table - 3] shows the results processed by the SPSS 9.05 software. The 5,7-dimethoxy-4-phenylcoumarin inhibited A427 cell growth by 42%, while 5,7-dimethoxy-4-p-methoxylphenylcoumarin inhibited it by 64% (FNx01 P < 0.05). These results suggest that 5,7-dimethoxy-4-p-methoxylphenylcoumarin is more active than 5,7-dimethoxy-4-phenylcoumarin inhibited, probably due to the methoxyl- group at the 4-carbon position on the phenyl of these compounds. The effects of 5,7-dimethoxy-4-p-methoxylphenylcoumarin, 5,7-dimethoxy-4-phenylcoumarin and ethanol solvent on bcl-2 and Bax oncoprotein expression after 24-h in the A427 cultures are shown in [Figure - 1]. Bcl-2 expression (survival action) was decreased in the A427 cells treated with 5,7-dimethoxy-4-p-methoxylphenylcoumarin and 5,7-dimethoxy-4-phenylcoumarin and Bax expression was slightly increased with these coumarins. Western blotting assays were repeated at least three times and a representative gel is included as [Figure - 1]. The densitometric values for the bcl-2, Bax and β -tubulin bands are given in [Table - 1]; the mean and SD of three normalized bcl-2 and Bax values with respect to β -tubulin (value = 1) are shown. Research in our laboratory indicates that the proliferation capacity of a human adenocarcinoma cell line (A427), overexpressing the bcl-2 oncogene, can be reduced when incubated with 5,7-dimethoxy-4-p-methoxylphenylcoumarin and 5,7-dimethoxy-4-phenylcoumarin. Using Western blotting, we have demonstrated with these cells that, when coumarins are present for 24-h in the cell culture medium, the bcl-2 protein is decreased, which raises the intriguing possibility that further intracellular growth signal processing is almost abandoned, leading to inhibition of cell proliferation. These observations are in agreement with the well-known antiproliferative effect of coumarins on malignant cell lines, decreasing bcl-2 expression and its survival action. There is an urgent need for agents with minimal side effects but with significant antitumor activity for particular cancers. 5,7-dimethoxy-4-phenylcoumarin might be effective in preventing or delaying the formation of metastases; they are candidates for adjuvant treatment strategies to delay metastases. Both 5,7-dimethoxy-4-p-methoxylphenylcoumarin and 5,7-dimethoxy-4-phenylcoumarin exhibit low toxicity in normal cells and their effect on malignant cells recommends them both as chemopreventatives and for use in combined antitumor treatment. Acknowledgement We are grateful to Prof. Pittaya Tantiwachwuttikul for isolation the pure compounds. This research project was funded by Thailand Research Fund MRG4980137 and Faculty of Science, Silpakorn University, Nakorn Pathom, Thailand.References
Copyright 2007 - Journal of Cancer Research and Therapeutics The following images related to this document are available:Photo images[cr07022t1.jpg] [cr07022f4.jpg] [cr07022t2.jpg] [cr07022f2.jpg] [cr07022f1.jpg] [cr07022f3.jpg] [cr07022t3.jpg] [cr07022t4.jpg] |
| |||||||||

{kind=link}
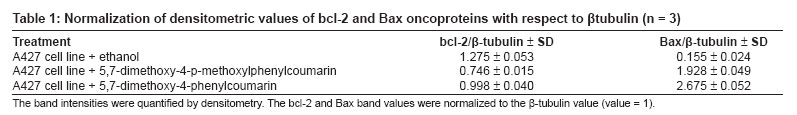{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}