 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
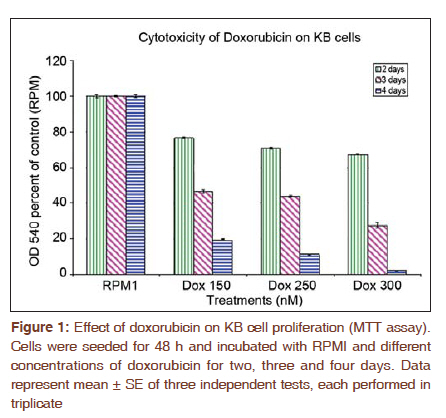
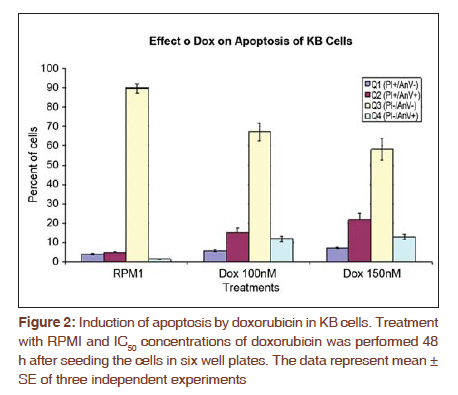
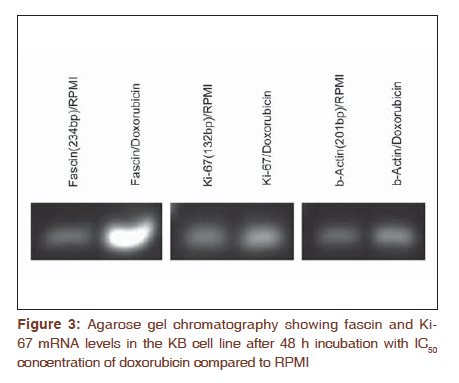
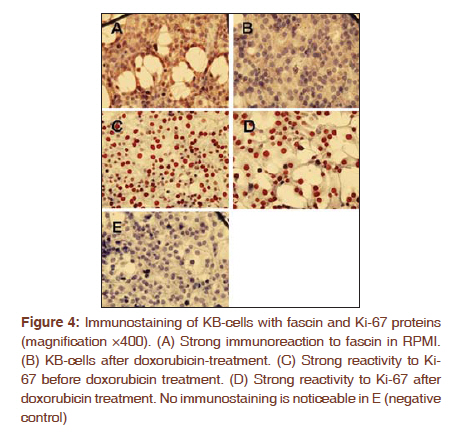
Journal of Cancer Research and Therapeutics, Vol. 7, No. 4, October-December, 2011, pp. 427-432 Original Article Expression of fascin protein and mRNA in the KB carcinoma cell line following treatment with doxorubicin Mojgan Alaeddini1, Shamileh Fouladdel2, Shahroo Etemad-Moghaddam1, Ebrahim Azizi3 1 Dental Research Center, Tehran University of Medical Sciences, Tehran, Iran Code Number: cr11115 DOI: 10.4103/0973-1482.92009 Abstract Context: Fascin is an actin-bundling protein that has been shown to be overexpressed in various carcinomas, but the effect of doxorubicin on its protein and mRNA has not been investigated. Keywords: Actin-bundling-protein, adriamycin, cancer, cell movement Introduction Cancer is a globally important disease with approximately 12.7 million new cases and 7.6 million deaths reported in 2008 and is estimated to affect 26 million people in 2030, causing 17 million deaths annually. [1],[2] Cancer mortality is mainly due to metastatic neoplasms that do not respond to routine treatments. [3] In order to metastasize, malignant cells undergo a series of complex events, during which they achieve significant motile properties leading to increased migratory abilities and invasion of surrounding tissues and vasculature. [3],[4],[5] Cellular movement requires redistribution of cytoskeletal actin into parallel bundles to provide additional strength for cell protrusions in the form of filopodia or lamellipodia. [3],[5],[6] These projections are regulated by a number of proteins like fascin. [3],[6] Actin is a principal cytoskeletal protein found in almost all eukaryotic cells. It functions in a variety of cellular processes including motility and is regulated by several other proteins. [7] Fascin is an actin bundling protein that primarily groups F-actin into well-organized parallel bundles via two actin-binding sites. [3] These assembled bundles constitute the major part of lamellipodia and filopodia required for cellular motility and consequently, metastasis. [6] Most nonpathologic differentiated epithelial cells are negative for fascin; however, this protein is overexpressed in a variety of carcinomatous tissues, especially in the invasive fronts [3],[6] and when the neoplasm is in a phase of "active invasion and metastasis". [6] In addition, a number of investigations have reported a relationship between fascin positivity and carcinomatous cell proliferation, with contrasting results. [3],[8],[9],[10],[11],[12] Doxorubicin (adriamycin) is a potent anthracycline chemotherapeutic agent that is known for its cytotoxicity and DNA-damaging effects. [13],[14],[15] Most studies conducted on different cancer types and cell lines have focused on the apoptosis-inducing effects of this anticancer drug. [14],[16],[17] However, a number of investigations have noticed additional qualities that are independent of its tumoricidal activity and have considered this agent to have a complex multilevel impact inside neoplastic cells. [18] For example, it has been suggested that doxorubicin can negatively influence the polymerization of actin in certain concentrations, resulting in reduced development of F-actin. [13],[19] Furthermore, this drug has been shown to exhibit antimigratory properties and the ability to increase focal cellular contact development. [18],[20],[21] The aim of the present study is to evaluate fascin in the KB cell line and investigate the possible changes induced by doxorubicin on its protein and mRNA expression. In addition, we studied these changes in light of alterations developed in Ki-67 following doxorubicin treatment. Materials and Methods All experiments were conducted in a triplicate manner and repeated at least three times. The KB human squamous cell carcinoma cell line (ATCC CCL-17, USA) was obtained and grown in monolayer culture in RPMI 1640 (BioSera, UK) supplemented with 10% fetal bovine serum (BioSera, UK) and penicillin-streptomycin (100 U/ml) at 37°C in a humidified CO 2 incubator containing 95% air and 5% carbon dioxide. [22] Cytotoxicity experiments were performed when KB cells reached 90% confluence in T25 flasks. The cells were trypsinized, washed with PBS and counted in a Neubauer slide using the trypan-blue dye-exclusion test. This was followed by seeding the cells into 96-well plates at a density of 5×10 3 cells/well for 48 h. The KB cells were then treated with 150, 250 and 300 nM doxorubicin. Control wells remained in the RPMI culture medium alone. Growth rate was determined by MTT assay after incubation for two, three, and four days as described previously. Briefly, each well was incubated with 4 mg/ml of MTT (25 μl) in PBS at 37°C in the CO 2 incubator. After 3 h, the medium and MTT were discarded from the wells and formazan crystals were solubilized in 100 μl of DMSO. Absorbance was recorded at a test wavelength of 540 nm (reference wavelength, 690 nm) using a microplate reader (Sunrise Tecan, Austria). [23] Results were expressed as mean ± SE and percent survival was calculated as the absorbance values of treated to untreated (control) wells. Six-well plates were seeded (5 × 10 5 cells/well) for 48 h and subsequently incubated with either control medium (RPMI) or doxorubicin at 50% growth inhibitory concentrations (IC 50 ) for another 48 h. In addition to IC 50 , doxorubicin was also used at a concentration of 100 nM. Media containing floating cells was also collected, washed with cold PBS and added to the cells harvested by trypsinization. Based on the instructions provided by the manufacturer, the cells were recollected and incubated with Annexin V-FITC and PI (APOPTEST TM -FITC kit, NEXINS RESEARCH, Netherlands), following centrifugation. Flowcytometric analysis was performed on the information acquired from 10 4 cells using a PARTEC flowcytometer equipped with FloMax software. [24] Reverse transcription-polymerase chain reaction (RT-PCR) was employed to determine the mRNA expression level of fascin and Ki-67. Briefly, KB cells were plated in T25 flasks and preserved in RPMI medium supplemented with FBS and pen-strep for two days. The IC50 of doxorubicin was added to the flask. After 48 h incubation, TriPure isolation reagent (Roche, Germany) was utilized to isolate total cellular RNA from untreated (control) and treated cells as previously described. [25] Total RNA (2 μg) was incubated with M-MLV reverse transcriptase (Fermentas) and oligo (dT) 18 for 1 h at 42°C followed by 10 min incubation at 94 °C in order to synthesize cDNA library. The Fascin and Ki-67 mRNA expression was determined by real-time PCR using 2 μl of prepared cDNA and 0.5 μl of each forward and reverse specific primers for fascin (forward 5'-CGCTCATCACCCTCGCCTTC-3'; reverse 5'-GCTCGTCCTTGCCCACCTTG-3') and Ki-67 (forward 5'-GAGAAAGAGTGGCAACCTGC-3'; reverse 5'-TCCGAAGCACCACTTCTTCT-3') at a concentration of 0.05 mM. β-actin was used as internal control (β-actin forward:5΄-TACTTGCGCTCAGGAGGAGCAA-3΄; reverse:5΄- GTCCTGTGGCATCCACGAAACT-3΄). In addition to the cDNA and primers, the qPCR reaction tubes contained 10 μl of Quantitect Syber Green Master mix (2x) and DEPC water to a final volume of 20 μl. The amplification program for fascin and Ki-67 consisted of predenaturation for 10 min at 95°C, followed by 40 cycles of denaturation (10s, at 95°C), annealing (10s, 60°C) and extension (10s, 72°C) and an ultimate cycle of final extension for 2 min at 72°C. The same procedure was carried out for β-actin, except that we used a temperature of 57°C during the annealing step. An agarose gel electrophoresis (1.5%) stained with ethidium bromide was also used to confirm single band of product with expected length for each amplified cDNAs. Analysis was performed by the ΔΔCT method, in which the relative mRNA expression of fascin and Ki-67 was normalized using β-actin expression. KB cells were incubated with RPMI 1640 culture medium in eight-well chamber slides (Lab Teck, USA) at 37°C in a humidified CO 2 incubator (5%) for 48 h, followed by exposure to the IC 50 concentration of doxorubicin for three days at 37°C. Control wells contained RPMI 1640, only. The cells were washed twice with cold PBS and fixed in methanol: acetone (9:1) for 30 min at -20°C. In order to block endogenous peroxidase activity and prevent nonspecific antibody binding, the fixed cells were placed in 3% H 2 O 2 and methanol for 30 min followed by Ultra V block (Labvision, USA) for 10 min. KB cells were then treated with primary mouse monoclonal antibody against fascin [SPM133 (ab15110); AbCam, UK] and rabbit monoclonal antibody against Ki-67 (SP6; LabVision, USA) at a dilution of 1:50 and 1:150 (overnight, 4°C), respectively. For antibody detection, a streptavidine-biotin immunoperoxidase detection kit was used with AEC as the chromogen (Labvision, USA) and necessary modifications to the manufacturer's instruction were made. Ultimately the cells were analyzed under a light microscope subsequent to counterstaining with Meyer's hematoxyline (DakoCytomation, Denmark). Negative controls consisted of cells in which the primary antibody was excluded. Fascin and Ki-67 expression were assessed based on the intensity of the stained cells, ranging from weak (+) to strong (+++). All data are the result of three tests in addition to a triple repetition for each condition and have been presented as means ± SE. Comparisons were performed using t-test and one-way analysis of variance (ANOVA) followed by Tukey's post hoc test to determine differences between groups (P < 0.05). Results KB cell cytotoxicity was assessed following doxorubicin treatment using the MTT assay. The antiproliferative effect of doxorubicin on KB cells was time- and dose-dependent [Figure - 1]. The 50% inhibitory concentration value of doxorubicin was calculated as 150 nM (three-days incubation) based on the results of the MTT test. Comparison of the effect of 48 h treatment with doxorubicin versus RPMI (control) on KB cells [Figure - 2] indicated a significant difference (P < 0.001) between the percentage of apoptotic cells detected by flowcytometry in Q4 (annexin V+/PI-). A significant difference (P < 0.001) was also observed between the control and treatment groups in Q2 and Q3. qRT-PCR was employed to determine fascin and Ki-67 gene expression in KB cells treated with doxorubicin. Agarose gel electrophoresis showed a single band with expected length for fascin, Ki-67 and β -actin [Figure - 3]. Real-time data for both fascin and Ki-67 mRNA expression in KB cells normalized to β-actin showed a significant increase (P < 0.001 and P = 0.049, respectively) in the presence of doxorubicin as compared to the RPMI control. Following immunostaining with fascin primary antibody, strong (+++) immunoreactivity was observed in the cytoplasm of KB cells. Doxorubicin treatment induced a substantial decrease in the cytoplasmic immunostaining of fascin (+) in comparison to RPMI control. Ki-67 immunoreactivity was located in the nuclei and did not show a considerable difference in staining intensity before and after treatment with doxorubicin [Figure - 4]. Discussion According to the results obtained in the present study, fascin protein and mRNA were expressed in cultured KB cells, a human squamous cell carcinoma cell line. Considering that normal squamous epithelium (skin and esophagus) is usually negative for fascin, except for its weak expression in the basal layer, [3],[26] we found an overexpression of this marker in our sample of epithelial malignant cells. Other investigations have also reported the overexpression of fascin in various cancer tissues and cell lines, some of which indicated a positive association with poor prognosis and aggressiveness. [4],[8],[27],[28],[29],[30] Comparison of different SCC cell lines has shown that tumors with a higher degree of aggressiveness, demonstrate stronger fascin expression. [5],[30] This could be explained by the cell-motility-promoting characteristics of fascin that leads to facilitation of cellular movement, required for invasion and metastasis. [3],[5] Fascin enhances motility through interference with actin filaments enabling them to produce cytoplasmic protuberances like filopodia and lamellipodia. [3],[6] Therefore, drugs that can inhibit or diminish the migration of fascin-expressing cancer cells via their interference with actin, may have a significant effect on the management of malignancies. Doxorubicin is generally employed for its cytotoxic, cytostatic and growth-inhibitory effects, [13],[14],[15],[16],[17],[18],[31] which were also observed in the present study. However, this drug has been shown to demonstrate other features including inhibition of invasion, [32] and metastasis, [33] increased development of focal contacts in melanoma cells, [18] and reduced motility.[18][19][20][21] The decrease in cellular migration can be explained based on the reported effects of doxorubicin on the actin protein. [19] Cellular actin can be found as either monomeric (globular actin or G-actin), or polymeric filamentous (F-actin) which exist in an equilibrium inside the cell. The dynamics of cytoskeletal actin which is known to be responsible for cellular functions like motility involves polymerization of G-actin into long filamentous actin and vice versa. [7],[34] Cell motility is achieved through forward protrusion of the membrane, following polymerization of actin filaments. [35] Doxorubicin has been shown to increase the cytoplasmic ratio of G-actin/F-actin, prevent polymerization of actin, inhibit filament growth and promote filament loss. [19] These drug-outcomes all seem to negatively affect the cellular requirements for actin-based cellular movement. Decrease in migratory features has been demonstrated in a human osteosarcoma cell line, [20] K1735-M2 mouse melanoma cells [18] and human HT1080 fibrosarcoma cells [36] following treatment with doxorubicin. All pharmaceutical agents that have any kind of impact on G-actin/F-actin polymerization are able to affect motility processes, [19] which may directly influence G-actin/F-actin transformation or act on actin-binding proteins. Considering the actin-bundling activity of fascin and that its protein decreased after doxorubicin treatment in the present study, it could be suggested that this actin-affecting drug may exert its anti-migratory effect by influencing fascin (possibly inhibiting it) in the KB cell line. This hypothesis requires further investigation with more sophisticated techniques and simultaneous examination of fascin, actin and doxorubicin. If this theory is proven to be acceptable, it might be useful in the development of anti-metastatic drugs either alone or in combination with other pharmaceutical agents like theanine [37] and taxol [20],[21] and may have clinical application pre-, peri- or post-chemotherapy. Furthermore, considering the increasing awareness regarding "personalized medicine", [3] fascin-positive neoplasms may benefit from doxorubicin or multidrug therapies and regimens which include doxorubicin as a constituting agent. In the current investigation, the effect of doxorubicin on fascin mRNA was also studied and a significant increase in fascin mRNA accompanied by a reduction in its protein was found using RT-PCR and immunocytochemistry, respectively. Despite the observation of fascin protein up-regulation in numerous carcinomas, the exact molecular mechanism underlying its increase is still unclear and could be transcriptional, post transcriptional or follow different routes in different cancer types. It has been suggested that various mechanisms may be responsible for the augmentation of fascin; however, transcriptional events or "alterations in mRNA stability" seem to have a more important role in some carcinomas. [3] In our study, a possible explanation for mRNA increase/protein decrease may be that doxorubicin inhibits actin polymerization, lowering migratory activity; consequently the carcinomatous cell attempts to increase its motility characteristics by up-regulating fascin m-RNA; however, post-transcriptional events, or other levels of dysregulation [3] possibly prevent the up-regulation of fascin protein. Doxorubicin also induced a similar increase in Ki-67 mRNA, which may be a significant finding regarding the association between fascin expression and proliferation, reported previously. [3],[8],[9] With respect to these findings, it might be suggested that fascin overexpression in KB cells may have an augmenting effect on Ki-67 mRNA. Hashimoto et al,[3] indicated that there may be increased amounts of phosphorylated fascin in carcinomatous cells, which by attaching to active protein kinase C (PKC) can lead to phosphorylation of cell-cycle-signalling molecules that ultimately result in cell proliferation. In the present study, the protein levels of Ki-67 did not change after doxorubicin treatment, which was in contrast to the decrease observed in fascin protein. As stated previously, we are not able to present an exact explanation for the mRNA/protein relationship found in this investigation; however, post-transcriptional events may have an important role in protein expression in the KB cell line. A considerable number of studies have examined the effects of doxorubicin in muscle and especially myocardial cells. It has been stated that due to the constant transformation of the aggregational state of actin in non-muscle cells and the fact that polymerization of G-actin is an important requisite for accurate cellular performances, doxorubicin damage could be a significant consideration in these cell populations. [19] However, the desirable effects of this drug on actin leading to reduced motility in non-muscle cells also influences cardiocytes and causes an unwanted decrease in contractile forces resulting in cardiotoxicity and disturbance in the overall functioning of the heart. [13],[19] Therefore, when planning a treatment regimen in which doxorubicin is included, its effect on muscle and non-muscle cells should be taken into consideration. To our knowledge, this is one of the first studies to examine fascin and doxorubicin simultaneously. Conclusion Considering the results obtained in the current study, it seems that this drug may exert a modifying effect on fascin protein and m-RNA, leading to possible applied impacts. Additional investigation using advanced methods and techniques are suggested to elucidate this hypothesis. References
Copyright 2011 - Journal of Cancer Research and Therapeutics The following images related to this document are available:Photo images[cr11115f3.jpg] [cr11115f2.jpg] [cr11115f4.jpg] [cr11115f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}