 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
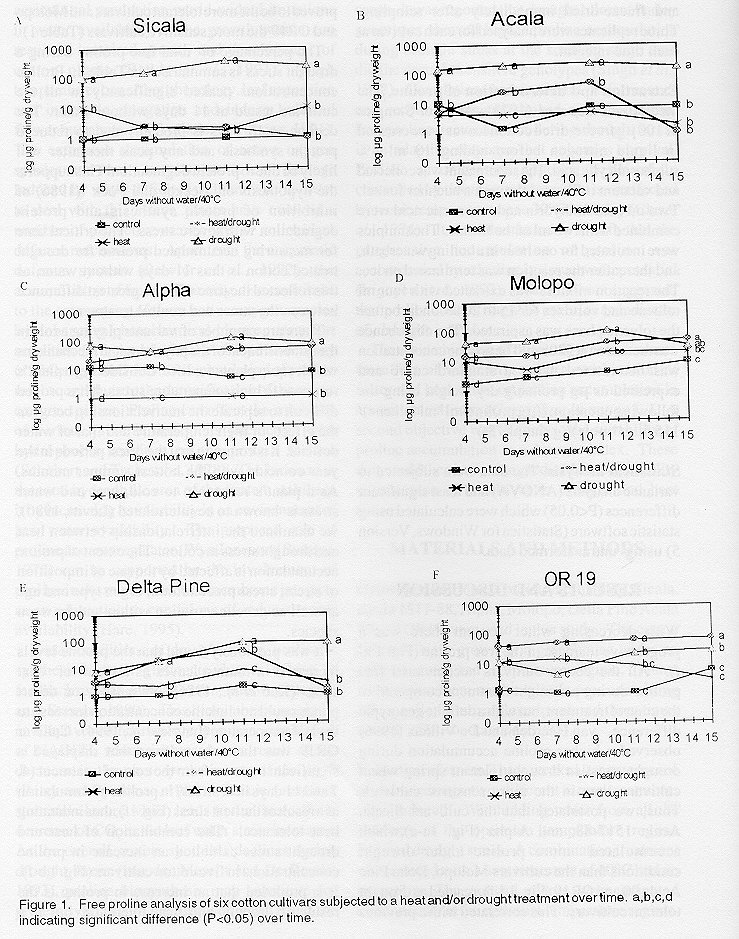
African Crop Science Journal, Vol. 8. No. 1, pp. 85- 91, 2000 SHORT COMMUNICATION PROLINE ACCUMULATION IN RESPONSE TO DROUGHT AND HEAT STRESS IN COTTON J.A. de Ronde, A. van der Mescht and H.S.F. Steyn ARC - Roodeplaat Vegetable and Ornamental Plant Institute, Private Bag X293, Pretoria, 0001, Republic of South Africa (Received 20 April, 1998; accepted 2 June, 1999) Code Number: CS00008 ABSTRACT Water and heat stress are the most important environmental variables affecting cotton growth and development. The main objective of our study was to evaluate the effect of water stress and a combination of water and heat stress on proline accumulation in six cotton cultivars (Gossypium hirsutum) and to determine the suitability of using this proline accumulation in a drought index. Drought and a combination of drought and heat stress were induced in three-week-old seedlings in the greenhouse. It was detected that with decreasing water content there was a progressive increase in free proline in all six cultivars, as well as differences in the proline level between the different cultivars. Maximum accumulation of free proline in drought stressed cotton occurred at 11 days without water. The combination of heat and drought stress exhibited an increase in proline concentrations in five cultivars. Different proline profiles were observed for the different treatments and different mechanisms for heat and drought are proposed for this phenomenon. Key Words: Drought stress, Gossypium hirsutum, proline RÉSUMÉ Les stresses hydrique et thermique sont les variables environnementaux les plus importants qui affectent la croissance et le développement du coton. L’objectif principal de notre étude était d’évaluer l’effet des stresses eau et une combinaison eau et chaleur, sur l’accumulation de proline dans six cultivars de coton (Gossypium hirsutum) et de déterminer la possibilité d’utiliser cette accumulation de proline dans un index de secheresse. La stresse hydrique et sa conbinaison avec la stresse thermique étaient induites chez les plantules de trois semaines d’âge en serre. Il a été détecté qu’avec une diminution du contenu hydrique, il y avait une augmentation progressive en proline libre dans les six cultivars, il en est de même pour les différences de niveau de proline entre les différents cultivars. L’accumulation maximum de proline libre dans le coton stressé de secheresse, est atteinte au 11me jour sans eau. La combinaison des stresses thermique et hydrique provoque une augmentation des concentrations en proline dans les cinq cultivars. Différents profils de proline étaient observés pour les différent traitements et différents méchanismes pour la chaleur et la sècheresse sont proposés pour ce phénoméne. Mots Clés: Stresse hydrique, Gossypium hirsutum, proline INTRODUCTION Drought and heat are the major causes of reduced agricultural production world-wide. Limitations in water availability in the growth season can result in impaired growth and development of cotton plants (Reddy et al., 1991). High temperatures can affect phenology, leaf expansion and internode elongation and induce boll and flower bud abortion (Trolinder and Shang, 1991). The extensive accumulation of active oxygen species and their contribution to cell damage induced by water deficit is well known. In order to deal with this effect, plants have evolved a number of protective scavenging or antioxidant defence mechanisms. Apart from the enzymatic defence system (Bowler et al., 1992), the accumulation of free proline may also contribute to the scavenging of these active oxygen species by enhancing photochemical electron transport activities (Alia et al., 1991). Floyd and Nagy (1984) suggested that the accumulation of proline might contribute to the detoxification of the active oxygen species. Proline can affect the solubility of various proteins due to its interaction with hydrophobic residues on the protein surface (Schobert and Tschesche, 1978). The increase in the total hydrophilic area of the protein stabilises it by increasing its solubility in an environment of low water availability. The proposed role of proline as osmoregulator (Wyn Jones and Storeys, 1978) can be supported by the involvement of proline in the maintenance of membrane integrity as an adaptation to conditions of reduced water availability (Hare, 1995). Under conditions of drought stress, proline is synthesised from glutamate due to a loss of feedback regulation in the proline pathway (Bogess and Stewart, 1980). This biosynthesis might be an adaptive mechanism to reduce the accumulation of NADPH, which increased as a result of the decrease in the photosynthetic rate of the plant (Berry and Bjorkman, 1980). This is a result of stomatal closure which leads to the intercellular decrease of CO2 as the leaf water stress increases. As the overall protein synthesis declines during drought stress (van der Mescht and De Ronde, 1993), proline biosynthesis may substitute for protein synthesis in the turnover of ATP and the oxidation of NADP (Hare, 1995). The rapid accumulation of free-proline in plant cells during drought stress is well documented for a number of crops. It was reported that drought tolerant barley genotypes showed higher proline accumulation during drought stress at the seedling stage than did the drought sensitive genotypes (Singh et al., 1972). Correlations between field performances and seedling tests encouraged speculation that free-proline accumulation during drought stress is an adaptive response that enhances survival. Proline accumulation appears to be solely controlled by tissue water status and is unaffected by tissue temperature up to 39°C in barley (Chu et al., 1974). It was also found in alfalfa that the progressive accumulation of proline was accompanied by a decline in tissue water potential (Irigoyen et al., 1992). These authors suggested that proline content might be useful as an indicator of drought-injury. The main objective of our study was to evaluate the effect of water stress and a combination of water and heat stress on proline accumulation in six cotton cultivars (Gossypium hirsutum L.). A second objective was to analyse the suitability of proline accumulation as a drought index. These data could then be used to predict the susceptibility or tolerance of a cultivar to drought and heat stress. MATERIALS AND METHODS Growth conditions. Six cultivars namely, Sicala, Acala 1517-88, Alpha, Molopo, Delta Pine Acala 90 and OR19 were used in this study. The cotton cultivars were grown in a greenhouse under an optimal water and temperature regime. Plants were grown from seed in plastic pots containing 2.5 kg soil mixture and were watered three times a week (De Ronde et al., 1995). Induction of stress. Three weeks after emergence of the seedlings two different stress treatments were induced. Withholding water from all the cultivars at the optimum temperature regime induced drought stress. A combination treatment was induced with a heat treatment of 40°C and no irrigation. The control plants were irrigated continuously at the optimum temperature regime. The leaf on the third apical node was harvested every five days from stressed and control plants. The leaves were quick frozen with liquid nitrogen and freeze-dried immediately after sampling. Three replicates were analysed for each cultivar at each time interval. Extraction and determination of proline. The method of Bates et al. (1973) was used. Samples of 100 µg freeze-dried cotton leaves were crushed in liquid nitrogen before adding 10 ml 3% sulphosalicylic acid. The supernatant was collected and vacuum infiltrated through a buchner funnel. Two ml acid ninhydrin and two acetic acid were combined with two ml of the filtrate. The samples were incubated for one hour in a boiling waterbath, and thereafter the reaction was terminated on ice. The reaction mixture was extracted with four ml toluene and vortexes for 15 to 20 seconds, before the toluene phase was aspirated. The absorbance was measured at 520 nm. The proline concentration was determined using a standard curve and expressed as µg proline/g dry weight using the following equation: ((µg proline/ml*ml toluene)/115.5µg/*mole)/(g sample/5)) Statistical analysis. The data were subjected to variance analysis (ANOVA) and least significant differences (P<0.05) which were calculated using statistic software (Statistica for Windows, Version 5) using untransformed data. RESULTS AND DISCUSSION Figure 1. Free proline analysis of six cotton cultivars subjected to a heat and/or drought treatment over time. a,b,c,d indicating significant difference (P<0.05) over time. With decreasing water content there was a progressive increase in the free proline (Fig. 1 a-f). All the cotton cultivars accumulated free proline during a drought treatment compared to the control treatment, but with a definite genotypic difference. van Heerden and De Villiers (1996) observed a higher proline accumulation during drought stress in drought tolerant spring wheat cultivars, than in the more sensitive cultivars. Thus, we postulated that the cultivars Sicala, Acala 1517-88, and Alpha (Fig. 1a-c),which accumulated more proline under drought conditions than the cultivars Molopo, Delta Pine Acala 90 and OR19 (Fig. 1d-f), reacted as drought tolerant cultivars. This correlated with a previous study (De Ronde and van der Mescht, 1997) using TTC viability assay as drought screening, where the cultivars Alpha and Acala 1517- 88 proved to be the more tolerant cultivars and Molopo and OR19 the more sensitive cultivars (Table 1) . Table 1. Comparison of sensitivities to (a) drought stress and (b) combined heat and drought stress in six cotton cultivars, as measured by proline accumulation and a TTC viability assay (De Ronde and Van der Mescht, 1997)
The percentage of total free proline during a drought stress is summarised in Table 2. Proline concentrations peaked significantly in all the cultivars tested at 11 days without water. The decline at 15 days is probably due to reduced protein synthesis and any peak thereafter will likely be due to protein degradation. This supports the hypothesis of Becker and Fock (1986) of inhibition of protein synthesis and protein degradation with severe stress. The critical time for measuring accumulated proline for drought treated cotton is thus 11 days without water, as this reflected the time with the greatest difference between the stress and control treatments. Table 2. The effect of drought stress on proline accumulation in six cotton cultivars (expressed as a percentage of the respective total proline accumulation)
z - indicating significant difference (P<0.05) There are a number of variants playing a role in the establishment of the physiological mechanisms which equip plants for life. Considering a plant’s response to high temperature stress, it has proved difficult to separate the interrelationship between the effects of high temperature and that of water deficits. It is common that the driest periods in the year coincide with the hottest summer months. As a plant’s resistance to cold, heat and water stress is known to be interrelated (Levitt, 1980), we examined the interrelationship between heat and drought stress in cotton. The extent of proline accumulation is affected by the rate of imposition of stress, stress precondition, organ type and age as well as genetic variation within and between species. It was previously found that the proline levels increased in tomato leaves as a result of heat stress (Kuo et al., 1986), but a study on desert plants could not link the concentration increase to the leaf temperature (Laurie et al., 1994). Cultivar OR19 was the only cultivar that displayed a significant increase from the control treatment (4, 7 and 11 days heat stress) in proline accumulation as a result of the heat stress (Fig. 1f), thus indicating heat tolerance. The combination of heat and drought stress exhibited an increase in proline concentrations in five cotton cultivars (Fig. 1 b-f). It is proposed that an increase in proline is the result of drought and heat tolerance, but if a drought tolerant cultivar accumulate less proline under high temperature, it is the result of heat sensitivity. This could be explained by possible different mechanisms for heat and drought and the activation of the heat stress mechanisms before the drought stress mechanisms. Thus, Alpha, Delta Pine Acala 90 and OR19 (Fig. 1 c, e and f) reacted as drought and heat tolerant cultivars, as they accumulated more proline compared to the cultivars Sicala, Acala 88 and Molopo (Fig. 1 a, b and d). This correlated with a previous study using TTC viability assay screening for combination of drought and heat stress (De Ronde and Van der Mescht, 1997), where the cultivars Alpha and Delta Pine Acala 90 proved to be the more tolerant cultivars and Molopo and Acala 88 the more sensitive cultivars (Table 1). Table 3 summarised the percentage of proline accumulation during a combined stress. It was observed that the critical stage for measuring proline is earlier with a combined stress than with a drought stress alone. Cultivars OR19 and Sicala showed a significant peak at 7 days without water. Cultivars Alpha, Molopo, Delta Pine Acala 90 and Acala 88 showed a significant peak at 11 days without water. Table 3. The effect of a combination of drought and heat stress on proline accumulation in six cotton cultivars (expressed as a percentage of the respective total proline accumulation)
z - indicating significant difference (P<0.05) CONCLUSIONS The data presented here indicate that there are differences in the proline level during a drought stress and those found during a combination of drought and heat stress. This is possibly a result of different mechanisms for drought and heat stress, with the heat stress mechanism firstly activated. It appears as if the tolerance or sensitivity of cotton cultivars can be differentiate with the use of a free proline analysis. We also observed that there are differences in the proline level between different cultivars. These results have potential for application in breeding programmes. ACKNOWLEDGEMENT We thank the Water Research Commission for their financial support. REFERENCES
©2000, African Crop Science Society The following images related to this document are available:Photo images[cs00008a.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}