 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
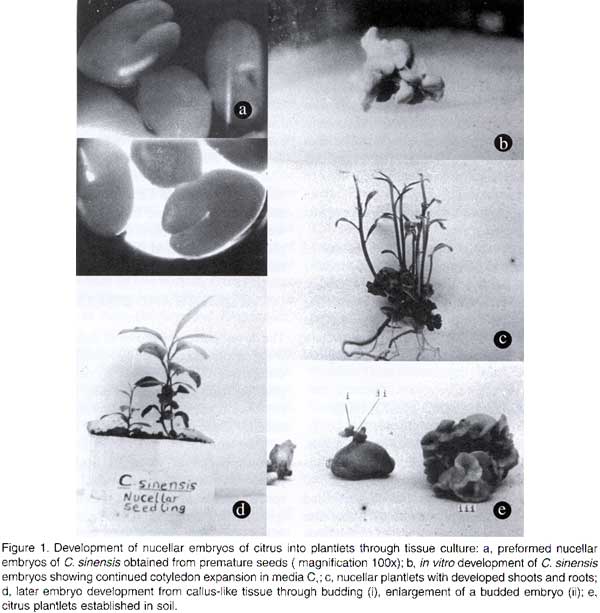
African Crop Science Journal, Vol. 8. No. 2, pp. 109-116 NUCELLAR EMBRYO CULTURE OF CITRUS SINENSIS L. AND CITRUS LIMON L. S. D. OBUKOSIA and KIMANI WAITHAKA (Received 5 September, 1997; accepted 10 May, 2000) Code Number: CS00011 INTRODUCTION Citrus production in East and Southern Africa is below 25% of the potential. Yields are low, ranging from 10 - 40 tonnes per hectare (Kolade and Olaniyan, 1998), while potential is up to 75 tonnes per hectare under high density planting (Wheaton et al., 1991). The low yields of citrus have been attributed to four major causes: the use of infected planting materials, use of non-budded planting materials, low use of fertilisers and irrigation, inadequate use of chemicals for pest and disease control, and planting of unimproved cultivars of scions and rootstocks. With regard to diseases, it has been observed that orchards of small-scale farmers are poorly managed. Many trees are usually infected with diseases, including the huanglongbing (HLB), citrus tristeza, cercospora leaf spot, scab and psorosis (Seif and Hillocks, 1993; Vuuren et al., 1999). Huanglongbing (HLB) previously called the citrus greening disease, is a severe and widespread disease of citrus in Eastern Asia (Pakistan to China), South and Eastern Africa, Arabic Peninsula, islands of Reunion and Mauritius (Bove and Garnier, 1992; Garnier and Bove, 1993). In Africa, the disease has been reported in Ethiopia, Kenya, Malagasy, South Africa, Sudan, Swaziland, Tanzania and Zimbabwe (Koizumi, 1995; Berg van den and van den Berg, 1999). HLB is caused by an uncultured phloem restricted bacterium that has recently been characterised (Jaguoiex et al., 1994). The ‘Candidatus’ generic name Liberobacter as defined by Murray and Shleifer (1994) for uncultured organisms has been given to the bacterium. The liberobacters from Asia and Africa belong to two different ‘Candidatus’ species, namely Liberobacter asiaticum and Liberobacter africanum (Villechanoux et al., 1993; Jaguoiex et al., 1994; Planet et al., 1995). HLB is transmitted among citrus trees by two vectors, Trioza erytrae Diaphorina (Del Guercio) in Africa and Diaphoria citri (Kaw) in Asia (Mclean and Oberholner, 1965; Capoor et al., 1967). It is difficult to get accurate estimates of yield losses from HLB disease in a perennial crop such as citrus. However, infected branches produce little or no marketable fruit. Experiments in South Africa to rejuvenate HLB infected citrus using tetracycline injection have reported increases in yields of up to 140% (Griesbach et al., 1979 unpubl.). In Kenya, yield losses of 100% have been reported. In order to exploit the production potential of citrus in Africa by reducing yield losses from diseases, there is an urgent need for farmers to plant seedlings obtained from registered nurseries which can guarantee cleanliness and trueness-to-type. Procurement of healthy planting material has been evasive in Kenya, where citrus from all registered nurseries, including the National Horticultural Research Centre, were found to be infected by the HLB or viral diseases or both. In Kenya, the sale and distribution of infected citrus planting materials from high altitude nurseries to low altitude regions accounted for over 90% of the geographical spread of HLB disease in the low altitude regions, which are climatically unfavourable to the vector Trioza erytrae. Thus, for African countries to realise increased and sustainable citrus production, two inter-ventions are required: nurseries should be supported by tissue culture methods for eradicating citrus diseases normally transmitted through planting material, and accurate molecular based disease diagnostic methods should be developed at the national level for seedling quality control. Three tissue culture techniques have been used to produce virus-free citrus plants: shoot-tip grafting, nucellar, and ovule cultures (Navarro, 1984). The later two approaches are effective because citrus viruses are not usually transmitted through seed, and nucellar plants produced through somatic embryogenesis have many characteristics similar to their mother plants. For example, Childs and Johnsons (1966) detected seed transmission of psorosis in Carizzo Citrange (a Poncirus x Citrus hybrid) but did not observe seed transmission of the virus within the genus Citrus. Bitters et al. (1970) reported that nucellar plants of Tempo tango, a monoembryonic hybrid (C. reticulata x C. sinensis), were free from exocortis, tristeza and veination diseases. Weathers and Calvan (1959) also reported on the importance of using nucellar embryony in polyembryonic citrus to obtain virus-free nucellar plants. Virus particles are generally restricted to the host’s vascular tissue, particularly the phloem. Since there is no direct vascular connection between the parent and either the zygotic or nucellar embryos, virus particles are eliminated from the seedling progenies (Button and Kochba, 1977). Rangan swamy (1958) first induced nucellar embryos from seeds (100-120 days post pollination) of an unspecified cultivar of Citrus species in in vitro culture, while Rangan et al. (1969) induced nucellar embryos from three cultivars of monoembryonic citrus. Polyembryony in citrus in vivo is controlled by multiple recessive genes (Maheshwari and Rangaswamy, 1958). In monoembryonic species, these recessive genes may synthesise a potent inhibitor of embryogenesis (Esan, 1973). Nucellar embryo culture has been successfully used in many monoembryonic cultivars of several citrus species, including C. grandis, C. aurantifolia x C. sinensis, C. clementine, C. reticulata and C. limon (cultivars: Ponderosa and Meyer lemons). However, the technique has not been successful with all monoembryonic clones (Bitters et al., 1970). Among polyembryonic citrus (with varying degrees of seediness), ovule culture rather than nucellar embryo culture has been used to induce nucellar plants in several cultivars (Bitters et al., 1970; Button and Borman, 1971; Button and Borman, 1974; Waithaka et al., 1988). Factors influencing successful regeneration of citrus from nucellar or ovule culture include genotype, age of the fruit at excision, presence or absence of fertilisation, and in vitro media composition (Bitters et al., 1970; Button and Kochba, 1977). Because of the importance of genotype in propagation systems (Cavalcante et al., 1994; Smith, 1997), it has been suggested that selection of responsive genotypes may lead to improved propagation capabilities and in regeneration of genetically transformed plants (Lowe et al., 1996). The current study investigated the response of two polyembryonic citrus species Citrus sinensis cv. Valencia Late and C. limon cv. Rough Lemon, to in vitro induction of adventive nucellar seedlings, with the objective of using the method to eradicate the HLB and viral diseases from nurseries and to propagate healthy seedlings en masse. MATERIALS AND METHODS Immature seeds of polyembryonic citrus cvs. Valencia Late and Rough Lemon were surface disinfected in 0.5% sodium hypochlorite for 10 minutes and rinsed three times in sterile distilled water. Zygotic embryos were excised with nucellar tissue, and during culture identified on the basis of their ability to develop rapidly into plantlets prior to being subcultured. Nucellar plants were identified based on their development from slow-growing proembryos which formed multiple cotyledons. Three media, C1, C2 and C3, were used to culture the nucellar embryoids. The three media contained Murashige and Skoog (1962) basal mineral nutrients plus sucrose (30 gl-1), myo-inositol (100 mg l-1), pyridoxine.HCl (1.0 mg l-1), thiamine.HCl (0.2 mg l-1), nicotinic acid (1.0 mg l-1), glycine (0.04 mg l-1) and Difco-Bacto-agar (10 g l-1).C3 contained all the above components alone. However, in addition to the above components, C1 medium contained 10% (v/v) filter-sterilised coconut water, while C2 contained 0.4 g l-1 filter-sterilised casein hydrolysate. The pH of the media was adjusted to 5.7 ± 0.1 using HCl or NaOH. The media were autoclaved at 121 °C at a pressure of 15 psi for 15 minutes. Disinfected nucellar tissues from Valencia Late and Rough Lemon seeds were aseptically excised and cultured in C1, C2 and C3 media. C3 medium was not used for continuous embryo initiation since it failed to support the growth of nucellar embryos. Enlarged embryos were subcultured every 30 days into fresh C1 orC2 media until they developed into plantlets with shoots and roots. The in vitro cultures were maintained under 18 hrs photoperiod at 28 ± 2.5 °C. In vitro produced nucellar plantlets were transplanted to soil and watered with MS mineral solution. Transplanted plantlets were covered with polythene sheets to maintain high relative humidity. RESULTS AND DISCUSSION The developmental patterns of nucellar plantlets from citrus in this study could be broadly categorised into two types: (i) development of plantlets from preformed nucellar embryos, and (ii) development of plantlets from callus-like nucellar tissue. Development of in vitro plantlets from preformed nucellar embryos. Both Citrus species possessed preformed nucellar embryoids (proembryos) and also exhibited later proliferation of nucellar embryoids from a white callus-like tissue (Fig. 1a). Similarly, Navarro et al. (1984) reported the presence of preformed embryos and later proliferation of embryoids from a callus-like tissue of citrus. Ranga swamy (1958) obtained nucellar embryos from an unspecified cultivar of Citrus species, and also noted that at the time of culturing the nucellar explants had already formed embryos in vivo. In the current study, C. sinensis L. cv. Valencia Late had up to 16 preformed nucellar embryos (Fig. 1a). These embryos were at different stages of development. The largest embryo, presumed to be zygotic, germinated precociously by developing into plantlets with shoots and roots within 30-60 days of in vitro culture. In contrast, nucellar embryos which were younger and smaller than zygotic embryos continued to develop in vitro through enlargement and formation of multiple cotyledons. Roots formed within 4 to 8 months while shoots took 6 to 9 months to develop (Fig. 1b) . Nucellar plants were maintained in vitro for 7 to 9 months from initiation until the multiple cotyledons degenerated (Fig. 1c). This dege-neration of multiple cotyledons prior to trans-planting to soil avoided rotting caused by secondary microbial infection. In contrast, Juarez et al. (1976) reported that 50% of the nucellar embryos of C. clementine cv. Clementine, with visible cotyledons germinated into plantlets that could be transplanted to soil within 1-2 months when sub-cultured into fresh medium. This was similar to the time taken for our zygotic embryos to reach transplanting stage. However, for the nucellar embryos, large embryos reached transplanting stage earlier than smaller embryos. The faster development of C. clementine cv. Clementine into plantlets (Juarez et al., 1976) compared to C. sinensis cv. Valencia Late is largely attributable to genetic differences between the two species. Embryos developed much faster into plantlets when separated into individual units than when left to grow together into clumps. Three developmental patterns of embryos in vitro were observed: some embryos developed into plantlets with multiple shoots and roots, others developed into structures with roots but no shoots, while a few embryos enlarged without differentiation into shoots or roots. One cause of development of embryos into plantlets with multiple shoots and roots was poor separation of embryos early in development, which led to the grafting of several plantlets. The other cause of multiple shoots was the growth of cotyledon buds. Development of one primary shoot with roots was attributed to apical dominance. Similarly, Chaturvedi and Mitra (1974) observed that callus-generated embryos of C. grandis when cultured in media with benzyl amino purine and naphthalene acetic acid produced 20-30 buds from the cut end and all developed into shoots. They explained that this was due to lack of apical dominance. Some C. sinensis cv. Valencia Late nucellar embryos which were left to grow together, produced no shoots or roots. However, the cotyledons continued to enlarge in clumps. This observation was similar to a report by Juarez et al. (1976), that embryos of C. clementine cv. Clementine rarely germinated if left attached to the callus or other embryos. Another observed abnormal growth pattern was the development of roots without shoots. It is possible that the later abnormality in development could be overcome by the addition of cytokinins into medium. Development of nucellar plants via callus tissue. Callus-like clumps formed in vitro from nucellus tissue of C. sinensis cv. Valencia Late and to a lesser extent in C. limon cv. Rough Lemon. In Valencia Late, a white callus-like tissue was observed after 4-5 months of in vitro culture. Callus formation had been reported by other investigators in ovule culture of C. sinensis cv. Washington navel (Navarro, 1984), nucellar culture of C. clementine cvs. Fina Clementine, other Clementine cvs. and Shamouti cv. of C. sinensis (Button and Borman, 1971; Juarez et al., 1976; Navarro, 1984). Light and electron microscope studies of Shamouti nucellar callus (Maheshwari and Ranganswamy, 1958) showed the presence of small globular proembryos or meristemoids (Torney, 1966), at various stages of development and absence of unorganised parenchymatous tissue. Embryogenesis occurred in single cells at the periphery or within existing proembryos. Proembryos may enlarge into spherical pseudobulbils of 4 mm (Button and Borman, 1974). However, in Shamouti cv. they rarely developed into plantlets. In the current study, however, when callus of Valencia Late cv. was subcultured into C1 or C2 media, the callus readily proliferated into embryoids with multiple cotyledons of about 1 to 3 mm in length within 19-27 days. The embryoids developed by budding with their radicles attached to a common spherical white tissue (Fig. 1d). Young embryoids (<1 mm long) left in the C1 or C2 medium without subculturing for up to 40 days turned brown and died. However, the callus tissue could continuously produce embryos by subculturing in fresh media each month. This was similar to an observation by Ranga swamy (1958) that nucellar tissue could be maintained in media supplemented with 0.4 g l-1 casein hydrolysate and kinetin (2 µg l-1) for over a year by continued subculturing. In the present study, embryos from callus subcultured at 30 days intervals developed into plantlets with roots within 6-10 months and some produced shoots after 8-11 months of in vitro culture. A few were transplanted to soil after 9.5 months of in vitro culture. Development of shoots and roots depended on the initial size of the embryoids at the time of in vitro culture. Medium to large embryoids (>2 mm diameter around cotyledon region) developed into plants with shoots and roots when subcultured in C1 or C3 medium. The development of callus into plantlets in C. sinensis cv. Valencia Late was generally slower than reported by Rangan swamy (1958). This difference could be attributed to either genetic differences between the Citrus species or differences in in vitro media used. Citrus limon cv. Rough Lemon formed limited white callus which rarely developed into embryos. This response was similar to the study of pseudobulbils of C. sinensis cv. Shamouti which rarely formed plantlets (Button and Borman, 1974). Also, Rough Lemon embryos did not develop multiple cotyledons as observed in Valencia Late. The only report of nucellar culture from C. limon was in two monoembryonic cvs. Ponderosa and Meyer lemon (Navarro, 1984). However, callus formation was not observed. The discrepancies between these two studies may be attributed to genetic differences between cvs. Ponderosa and Meyer lemon which are monoembryonic, while cv. Rough Lemon is polyembryonic. It has been reported that monoembryonic citrus synthesise a potent anti-embryonic substance that could be a major limiting factor in the initiation of nucellar embryos in vitro (Esan, 1973). Establishment of plants in soil. Normal plantlets, with well developed roots and shoots and degenerated cotyledons were transplanted to soil (Table 1, Fig. 1 e). Significantly more Rough Lemon plantlets were established in soil than Valencia Late (Table 1). Plantlets that failed to establish in soil were first stunted in growth and later decayed. Victor and Arambulla (1984) reported that transplantation to soil was a critical stage of tissue culture and this step is often unsuccessful. In our study, plants transplanted to soil prior to the degeneration of the cotyledons died from rotting promoted by decomposing cotyledons. It is also possible that the intertwining nature of multiple cotyledons made it hard to completely wash-off all agar from the plants hence leading to secondary infection. We therefore emphasise that plants should be maintained in vitro until the cotyledons degenerate. Perhaps this prolonged in vitro culture also ensures development of the vascular connection between the shoots and roots, and also the development of an optimum proportion of root to shoot systems as suggested by Sommer and Caldas (1981).
Often the plants were observed to die due to microbial infection from the soil. Use of sterile soil greatly reduced this problem. Transplanted plants were also very susceptible to attack by white fly larvae. The soft bark tissue was easily eaten around the crown resulting in death. This problem was checked by applying insecticides to control white flies. CONCLUSIONS In this study nucellar plants were successfully produced from Valencia Late and Rough Lemon cultivars of citrus emanating from both preformed nucellar embryos and from embryoids which budded on pseudobulbils produced from callus. Preformed embryoids developed into plantlets faster than pseudobulbils. The production of plantlets was very prolific, and therefore offers great potential for eradication of the greening and viral diseases and could also be used for rapid mass production of citrus rootstocks, especially the Citrus limon currently used as a rootstock in Kenya. The plants produced showed no signs of greening and viral diseases, although the starting materials were heavily infected. The tissue culture protocols developed in this study are currently being used to produce HLB-free seedlings for citrus farmers in Kenya. REFERENCES
The following images related to this document are available:Photo images[cs00011a.jpg] | |||||||||||||||||
| |||||||||

{kind=link}