 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
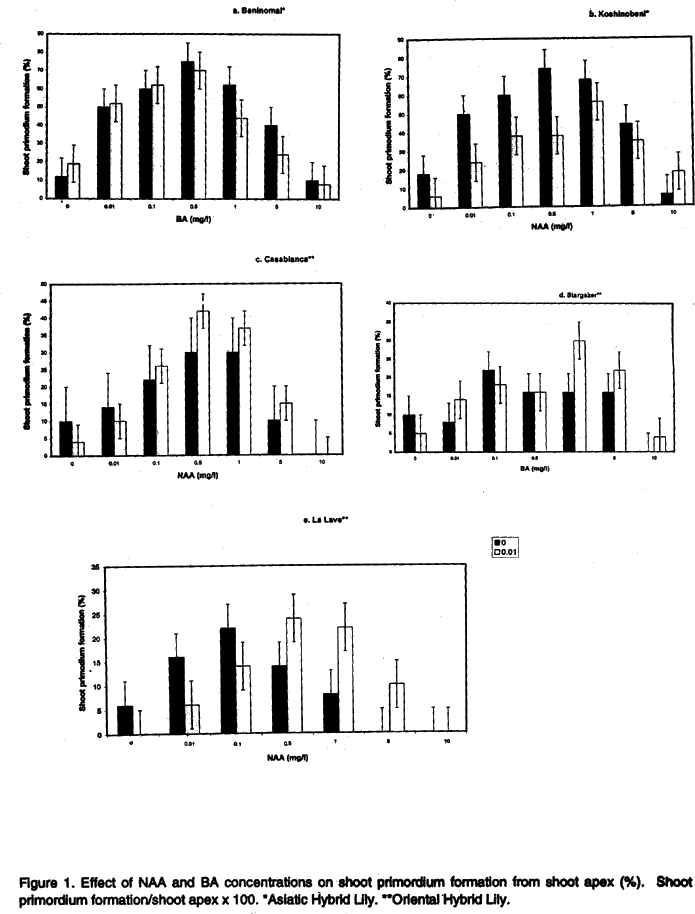
African Crop Science Journal, Vol. 8. No. 2, pp. 117-127 Comparison between Embryoid and Shoot Primordia Methods of Plantlet Production for‘Asiatic Hybrid Lily’and‘Oriental Hybrid Lily’ H. Sugiura, S.G. Agong, A. Enami1, H. Kaneko and T. Honma2 (Received 19 October, 1999; accepted 14 June, 2000) Code Number: CS00012 INTRODUCTION Production of lily bulbs plays a vital role in the cut flower industry and has been steadily increasing over the recent past within Kenya. Most materials used for propagating the lilies in Kenya are temperate (Northern Hemisphere) in origin. However, expansion of the lily bulb production has met severe restriction following virus infection outbreak in the crop (Malmd, 1997). Consequently, there is an urgent need to save the lily bulb production in the country from getting ruined. The livelihood of many small-scale growers is dependent on the lily bulb production, thus necessitating continuous availability of disease-free propagation material for their use. Furthermore, the country relies heavily on horticultural export (of which lily bulbs are considerably important) for foreign exchange. Among the propagation techniques, tissue culture is a useful technique for macro-propagation of virus-free lily bulbs (Sugiura, 1994). This technique could help in averting the virus infection of lily bulbs in Kenya and ensure production of disease-free planting materials. However, the ability of the Asiatic Hybrid Lily (Lilium x elegance)’ and the Oriental Hybrid Lily (L. spp. hybrid) to propagate via tissue culture is much less compared to L. longiflorum. This imposes a higher commercial price for the former two types of lilies in both local and external markets. Increasingly, only a few small-scale growers can afford the planting materials for the lilies. Bulbs of the Oriental Hybrid Lily are the most expensive in hybrid category followed by those of the Asiatic one. Because these hybrid lilies are the most preferred in the market, it is important to develop alternative procedures for their propagation with the aim of reducing the cost of production. Young bulbs of L. longiflorum propagated from the sucker can flower after one year of culture. However, young bulbs of the Oriental Hybrid Lily propagated from such suckers fail to flower within one year of propagation, and require at least two years of growth before the bulbs are ready for cut flower production. On the other hand, the commercial price of bulbs of L. longiflorum has been significantly reduced in the recent past via macro-propagation (Takahashi et al., 1991; Tanaka et al., 1991) whereas prices of the Oriental Hybrid Lily and the Asiatic Hybrid Lily bulbs are still relatively high because they cannot be macro-propagated. Furthermore, ones used for cut flower production, bulbs of the Oriental Hybrid Lily and the Asiatic Hybrid Lily are difficult to re-use continuously in the following year without replacement. This is due to poor cut flower quality arising from inability to support plant growth to the required height. Normally, bulbs selected from a newly bred lily variety are subjected to macro-propagation, and ten to twenty suckers can be obtained from one lily bulb when the suckers are used for propagation (Niimi, 1984). Thus, macro-propagation of lily bulbs remains important in the production of lilies. In an attempt to ensure continuous supply of lily plantlets, in vitro culture has been employed with varying success rates depending on varieties (Simmond and Cumming, 1976). In other trials, Niimi (1986) successfully propagated six lily varieties from leaf segments by tissue culture. Bulbs of L. longiflorum (Takahashi et al., 1991) and those of the Oriental Hybrid Lily (Hachi et al., 1992) have also been obtained via sucker culture in a liquid medium. Sucker culture systems result in rapid growth as well as production of many explants required for macro-propagation. Use of shoot primordia and embryoid techniques have also been advocated for macro-propagation of the lilies (Tanaka et al., 1991; Sugiura, 1994). A shoot primordium culture gives rise to virus-free tissue given that it is obtained from the apical meristem tip. Successful shoot primordia production in L. longiflorum via apical meristem tip culture has been reported (Tanaka et al., 1991). Shoot primordia production of the Oriental Hybrid Lily and the Asiatic Hybrid Lily from the meristem tip via protoplasts culture has also been reported (Sugiura, 1993). Also, embryoids do offer an alternative for lily propagation, especially given relatively lower labour costs compared to that for the shoot primordia. Minimal protocol is required for the embryoids techniques thus eliminating the bulk of labour cost in propagation (Sugiura, 1994). Embryoids are not only suitable for long term storage in super-freezers (Sakai, 1991), with no regeneration difficulties, but are also easy to transport in the from of artificial seeds (Murashige, 1978; Yakuwa and Oka, 1987). Nonetheless, an important drawback in plants regenerated from tissue culture is accompanying mutations that occur with a consequence of gradual deterioration of genetic purity of the crop. We therefore focused on possible somaclonal mutations arising out of embrioids and shoot primordia, and assessed their commercial viability as potential systems for lily propagation. MATERIALS AND METHODS Shoot primordium formation. The shoot primordia of the Asiatic Hybrid Lily varieties, ‘Beninomai (L.x elegance var.Beninomai)’ and ‘Koshinobeni (L.xelegance var. Koshinobeni)’, and also the Oriental Hybrid Lily varieties, ‘Casablanca (L. spp. hybrid var. Casablanca )’, ‘Stargazer (L. spp. hybrid var.Stargazer)’ and ‘La Lave (L. spp. hybrid var. La Lave)’ were obtained from apical meristems. Apical meristem tips were cultured in liquid medium using a 2 rpm inclined rotary shaker for two months according to the procedures of Tanaka et al. (1991), Sugiura (1993) and Sugiura et al. (1994). The liquid medium was a modification (1/2 major basal element and 0.7 g agarose) of that used by Murashige and Skoog (1962), with the concentrations of the hormones used as outlined in Figure 1. Regeneration tissues were removed from shoot promordia one month after culture. The propagated shoot primordium was divided in the piece followed by culturing lasting a few months. Regeneration from shoot primordium. The shoot primordia between 2 mm and 5 mm were planted in a solid medium. The regeneration medium used in all experiments was a modification (1/2 major elements plus 0.7 g agarose) of that used by Murashige and Skoog (1962), with the concentrations of hormones as given in Table 1. The number of plants generated from each shoot primordium was determined two months after planting.
Embryoid production. ‘Beninomai’ and ‘Benisugata’ Asiatic Hybrid Lily, and ‘Casablanca’, ‘La Lave’ and ‘Stargazer’ Oriental Hybrid Lily were used in the study. Embryogenic calli for producing embryoids were obtained by cutting root tips, the lower part of young bulbs and the suckers from the cut circumference of in vitro bulbs. They were cultured in the same medium as that for the shoot primordium, and the calli were formed after two months. The calli were strained as described by Otsuki et al. (1988). The size of the mesh was 2 mm, and the separated cells were cultured at 90 rpm using a level rotary shaker in a liquid medium supplemented with naphthalene acetic acid (NAA) and 6-benzylaminopurin (BA) (Table 2). Young plants and embryoid formation were investigated after three weeks.
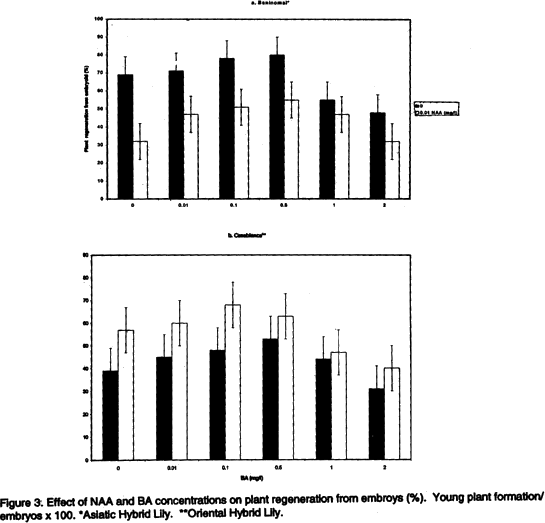
Regeneration from embryoids. Beninomai and Casablanca embryoids were planted in the regeneration medium to ensure smooth development. Plant regeneration was investigated after two months. Acclimatisation of the in vitro plants. ‘Beninomai’ plantlets were subjected to acclimatisation experiments. The plants were transplanted in fresh regeneration medium to allow for growth of the small bulbs (Sugiura et al., 1994). Bulbs over 6 mm grown in vitro from shoot primordia and embryoids were stored in vermiculite at temperatures of between 2 to 5 °C (refrigeration temperature) for two weeks in non-sealed plastic bags, and were covered with Benlate-T to effect disinfection during the transfer in vivo. The young plants were planted in nursery after cold storage in beds comprising of soil and farmyard manure. Later, the bulbs were planted in nursery beds or in pots under mild humidity, after cold storage followed by assessment of the proportion of bulb that decayed two months later. Mutation occurrence. Beninomai’, ‘Koshi-nobeni’, ‘Casablanca’, ‘Stargazer’ and ‘La Lave’ were used to study mutation rates. The young bulbs from in vitro suckers, shoot primordia and embryoids were planted in a nursery in vivo two weeks later, following cold storage in a refrigerator. The mutant- regenerated plants were recognised via tissue culture, in vivo occurrence of albino, and typical variations of young plants. RESULTS Shoot primordium formation. The highest proportions of shoot primordia of both Asiatic Hybrid lilies was found in 0.01 mg/l NAA and 0.5 mg l-1 BA medium (Fig. 1 a and b: non-significant), and for the three Oriental Hybrid lilies in 0.01 mg l-1 NAA and 1.0 mg l-1 BA medium (Fig. 1 c,d and e: non-significant). The shoot primordia of the Asiatic Hybrid lilies formed a membraneous structure with a cream colour unlike the case of the Oriental Hybrid lilies which formed a membranous-like embryogenic calli and had a transparent green colour (Fig. 2 a). The plantlets were transplanted after three months as they were fully grown in the test tubes. Noticeably, the shoot primordium showed brown colour as the sub-division by a scalpel and tweezers for transplantation progressed. Therefore, a fragment of the shoot primordium was placed into new medium in the subsequent multiplications. Regeneration from shoot primordium. Plant regeneration from shoot primordium occurred soon after planting in the regeneration medium. For ‘Beninomai’, a concentration of 0.01 mg l-1 NAA and 0.5 mg l-1 BA, respectively, produced the largest number of plants (Table 1). For ‘Casablanca’ the concentrations of 0.01 mg l-1 NAA and 0.1 mg l-1 BA were suitable, but the variant regenerated very small plants (Table 1, Fig. 2 b). Embryoid production. The highest ratio of embryoid was formed by the lower part of the apical meristem of young bulbs. In all lilies, the embryoids were scarcely formed, and their tissues were brown when less than 2 mm size mesh was used for isolation of embriogenic callus (Fig. 2 c). Numerous torpid embryoids and small embryogenic calli formed easily with a 2 mm mesh. It was difficult to discriminate embryoids from small embriogenic calli. Therefore, our data also comprised of the smaller embryogenic callus formation. The highest number of embryoids in ‘Beninomai’ was formed with concentrations of 0.01 mg l-1 NAA and 0.5 mg l-1 BA, while the highest number in ‘Casablanca’ formed with 0.01 mg l-1 NAA and 0.1 mg l-1 BA (Table 2, Fig. 2 d). Substantial differences amongst the treatments were observed and the trend for shoot primordia formation was very consistent among all the lilies (Fig. 1 a and c). Regeneration from embryoids. Plant regeneration from embryoids began soon after placement in the regeneration medium. The number of regenerated plants from shoot primordia differed between ‘Beninomai’ and ‘Casablanca’. The largest number of plants of ‘Beninomai’ was produced with a concentration of 0.01 mg l-1 NAA and 0.1 mg l-1 BA (Fig. 3 a), and for ‘Casablanca’ with a concentration of 0.01 mg l-1 NAA and 0.5 mg l-1 BA (Fig. 2 e, Fig. 3 b). Plants regenerated from the shoot primordia sometimes had two to three embryos. The bulbs therefore had to be separated when two to three bulbs grew from an embryoid. In some cases, regenerated and separated calli were primordia and not embryoids because the latter exhibited smaller calli due to the size of the mesh used. Acclimatisation of the in vitro plants. Sealing with a plastic wrap increased the humidity under in vitro lily bulbs by 3-8 %, unlike the case of the unsealed young plants of ‘Beninomai’. Liquid fertiliser was reduced by 10-20 % for in vitro lily bulbs acclimatisation treatment to prevent the bulbs from decaying. Longer cold storage of young lily bulbs before planting was found necessary to allow for collection of the regenerated bulbs (Table 3). The surviving bulbs of ‘Casablanca’ regenerated from shoot primordia and embryoids produced some leaves 1-2 months after planting in the nursery without maintenance of the humidity (Fig. 2 f).
Mutation occurrence. The albinos of ‘Beninomai’, ‘Casablanca’, ‘Stargazer’ and ‘La Lave’ occurred at the rate of 1 per 120-200 bulbs from shoot primordia and embryoids when cultured in vitro. Most albinos of ‘Casablanca’ occurred by regeneration from shoot primordia (Fig. 4 a). No albinos occurred by regenerating from suckers in any of the varieties studied (Table 4). Albinos were easily removed because of their early detection. Variation arising from albino formation was very rare in the field. In ‘Beninomai’, variation occurred at the rate of 1 per 1,500 plants and appeared in petals (Fig. 4 b). However, no mutational variations were detected in the chromosomes of ‘Beninomai’ (Fig. 4 c).
DISCUSSION NAA and BA requirement for both shoot primordium and embryoigenic callus formation, along with the regeneration in the medium in the case of the Asiatic Hybrid Lily tissue, were different from those of the Oriental Hybrid Lily tissue (Tables 1 and 2, Figs. 1 and 3). Also, in each variety, the NAA and BA requirement for embryoid formation and regeneration of the lilies were similar to those for shoot primordium formation and regeneration. Thus, these results are in full conformity to the observations of Sugiura (1993). Therefore, the hormone requirement for the lily tissue is guided by the family and has no varietal specificity. We believe that the NAA and BA requirements in the medium for the lilies were similar because the shoot primordia or the embryoids were regenerated from callus under the same conditions. As would be expected, embryogenic calli are formed by cutting the root tip of in vitro bulbs (Mii et al., 1991) and from the sucker of the cut circumference of in vitro bulbs (Sugiura, 1993). In this study, embryogenic calli from the lower part of young bulbs were superior for embryoid formation because the tissue near the apical meristem tip showed vigorous cell division. Most tissue from embryogenic calli were dead with only a few surviving, and following filtration through a mesh smaller than 2 mm possibly due to the injury caused on the tissue of the embryogenic calli. Shoot primordia of the lilies were transplanted in a fresh liquid medium because they had grown to full size in the test tube. Whenever calli were formed from shoot primordia in regenerated lily plant, they had to be separated from the plant each time shoot primordia were used to produce seedlings. Therefore, lily seedlings production from shoot primordia is a labour-intensive and thus very costly exercise. However, Tanaka et al. (1991) showed that more young plants are regenerated from the shoot primordium in L. longiflorum. On this basis, it is possible to obtain greater returns in terms of plantlet production in the Asiatic Hybrid Lily and the Oriental Hybrid Lily. Nevertheless, L.longiflorum has a greater capacity to propagate than the Asiatic Hybrid Lily and the Oriental Hybrid Lily. Sometimes embryogenic calli were formed from embryoids during the regeneration of plants. Lily embryoids can therefore be repeatedly produced from embryogenic calli by embryoid regeneration. Commonly, maintenance of humidity is necessary for the production of seedlings obtained from tissue cultures. Most of the young plants would wilt in the event that humidity is not maintained, given that they readily dry up through loss of water. However, young lily plants wither instead of wilting because of the bulbs. Also, high storage temperatures following cold storage of bulbs in L.longiflorum result into faster growth of the plants (Dennis et al.,1981 a, b), suggesting further that there is no need for extra energy in the hardening execise. Our method has a greater commercial potential than currently available for lily propagation. Moreover, only insignificant amount of soma clonal variations occurred. High mutation rates would make a procedure like the one described here totally unacceptable for commercial venture. For example, Larkin and Seoncrofe (1981) and Evans and Sharp (1983) have reported 2-20% occurrence of mutations in tomato and carrot plants regenerated from tissue culture. This is relatively high mutation rate and would not be suitable for a commercial system where consistency, top genetic and cut flower quality are critical criteria for guaranteeing the market. In this study, we did not experience such difficulties with the propagation of lily using shoot primordia or the embryoids given that only an insignificant amount of mutation occurred. Our method is useful for commercial production of the Asiatic Hybrid Lily as well as the Oriental Hybrid Lily seedlings. Consistently high quality of these crops would fetch premium price in the markets than L.longiflorum. It is particularly useful for ‘Casablanca’ embryoids due to its extremely expensive bulbs arising from exorbitant costs of propagation. REFERENCES
The following images related to this document are available:Photo images[cs00012b.jpg] [cs00012d.jpg]Line drawing images[cs00012a.gif] [cs00012c.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}