 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 8. No. 2, pp. 187-193 TWO BARLEY YELLOW DWARF LUTEOVIRUS SEROTYPES ASSOCIATED WITH STUNTING OF IRRIGATED WHEAT IN ZAMBIA J. NDUNGURU* and R. G. KAPOORIA1 (Received 1 March, 1999; accepted 3 March, 2000) Code Number: CS00020 INTRODUCTION Barley yellow dwarf virus (BYDV), the type member of the luteovirus (Metthews, 1982), comprises a group of serotypes or strains that infect barley (Hordeum vulgare L.), oats (Avena sativa L.), wheat (Triticum aestivum L.), rice (Oryza sativa L.) and other graminaceous hosts (Rochow, 1970; Wen and Lister, 1991). Five serotypes of BYDV were described by Rochow (1970) from New York (NY), namely NY-PAV, NY-MAV, NY-SGV, NY-RPV and NY-RMV. The five serotypes, designated PAV, MAV, SGV, RPV and RMV (Casper, 1988), were named after their specific vector aphid species, with vector specificity approximately corresponding to serotype (Rochow, 1970). The major vectors for the serotypes (in parentheses) are: Rhopalosiphum padi (RPV), Rhopalosiphum maidis (RMV), Sitobion avenae (MAV), Schizaphis graminum (SGV), and R. padi, S. avenae, and others (PAV) (Miller and Rasochova, 1997). BYDV can be separated into two subgroups, based on serological relationships that are presumably governed by viral caspid protein antigenicity (Vicent et al., 1990), cytopathology (Gill and Chong, 1979) and dsRNA ‘fingerprints’ (Gildow et al., 1983). Subgroup I includes PAV, MAV and SGV and subgroup II RPV and RMV (Rochow, 1970; Waterhouse et al., 1988). Barley yellow dwarf virus causes the most widespread and economically important virus disease of cereals (Rochow, 1970; Plumb, 1983; Wen and Lister, 1991), and can cause significant economic losses in wheat (Burnett, 1984; Waterhouse et al., 1988). Herera and Quiroz (1984) have estimated yield losses of between 10% and 60% caused by BYDV in wheat. In north Africa, BYDV causes serious losses in wheat when infection occurs early in the growing season (Comeau and Makkouk, 1992). The virus causes chlorosis and mosaic on infected wheat leaves (Brunt et al., 1990), and if infection occurs in the seedling stage plants appear stunted and may be half to one third their normal size (Wiese, 1987). Depending on the particular strain and its virulence, BYDV infection may inhibit root growth, reduce or prevent flower production, or increase host susceptibility to opportunistic pathogens, drought and other stresses (Burnett, 1984). Barley yellow dwarf disease has been diagnosed historically first by visual assessments and later by insect transmission tests, and most recently by enzyme-linked immunosorbent assay (ELISA) (D’Arcy et al., 1989), immunosorbent electron microscope (ISEM) (Forde, 1989) and polymerase chain reaction (PCR) (Robertson et al., 1991). Luteovirus detection and identification by ELISA has become routine, can be done on a large scale (D’Arcy et al., 1989) and can distinguish the various serotypes (Irwin and Thresh, 1992; Figueira et al., 1997). Dot immunobinding assay, a form of enzyme immunoassays having no antibody capture step, do not work reliably with viruses present in low concentration such as BYDV (Torrance, 1998). ELISA using polyclonal antisera is the preferred technique (Lister and Rochow, 1979). This paper presents the first report on the serological detection of BYDV serotypes and disease incidence in irrigated wheat in Zambia. MATERIALS AND METHODS Virus sources. Irrigated commercial wheat fields in Central, Lusaka and Southern provinces of Zambia were surveyed during June to August 1997 for the occurrence of BYDV. A total of 9 wheat cultivars, namely Deka, Gamtoos, Loerie II, MM2, Nata, Nkwazi, P7, Scan, and Sceptre, were inspected in 11 wheat fields. Sixty to seventy percent of each field was inspected following a walking pattern of Feistritzer (1975). Plants were examined along the line, and a few in each field of those showing suspected virus symptoms sampled by removing young symptomatic leaves (8 to 12 per plant) after describing the symptoms. Leaves from one plant constituted a sample. Some colour print photographs were taken for comparisons with those in the literature. Collected samples were put in plastic bags, sealed, labeled and transported in a cool box to the University of Zambia. Samples were kept at -20 °C and analysed within 7 days of storage. A total of 68 samples were collected in this study (Table 1).
Diagnostic technique. The samples were analysed with a direct double antibody sandwich ELISA (DAS ELISA) as described by Clark and Adams (1977), using reagent sets for testing BYDV serotypes (PAV and RPV) procured from Agdia Inc. (U.S.). The two serotypes were chosen to allow detection of BYDV isolates belonging to either subgroup of BYDV. As mentioned above, serotypes PAV and RPV belong to subgroup I and II, respectively (Aapola and Rochow, 1971; Rochow, 1970, 1979a). ELISA was done following the manufacturer’s (Agdia) instruction. Reconstituted lyophilised positive controls (leaf materials containing BYDV serotypes PAV and RPV from Agdia Inc.) were included. Sap from healthy plant leaves was used as negative control. ELISA reactions were evaluated by visual assessment of colour changes, and were recorded as follows: - no colour change (negative results), + pale yellow, ++ bright yellow, and +++ very bright yellow (positive results). Disease incidence. Within each field, a 1 x 1 m quadrant was selected along the transect line at every 10 m. The number of diseased plants and total number of plants in each quadrant were counted. The proportion of infected plants was then calculated using a statistical formula of Sammons and Barnett (1989) and expressed as percentage. RESULTS Disease symptoms. Wheat plants infected with BYDV serotypes PAV and RPV expressed mild to very severe symptoms (Table 2). The infected plants were markedly stunted with excessive tillers in some cases, which were very short. Severe symptoms consisted of bright yellow leaf mosaic beginning from the tips towards the base. Where both BYDV-PAV and BYDV-RPV co-infected, they induced very severe symptoms (Table 2). On wheat cultivar Loerie II, infected with serotype PAV, plants had distorted, curled and serrated leaves. On some fields plant leaves were narrow and stiff and tillers were completely suppressed. At Golden Valley Agricultural Research Trust (GART), where wheat cultivars Deka, MM2, P7, Nata and Sceptre had been planted on the same date, symptoms of BYDV were more severe on cultivars P7 and Nata than on the other cultivars.
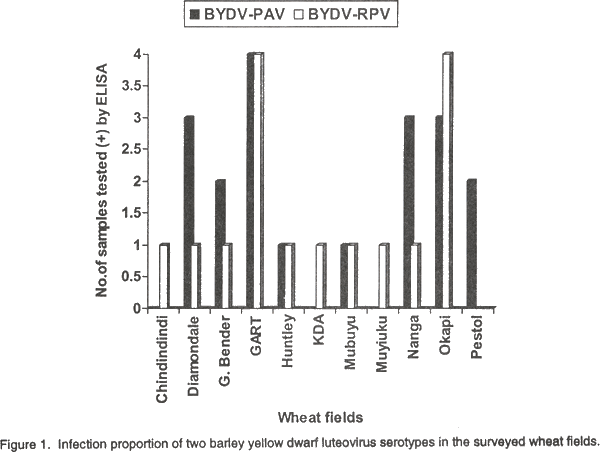
Serology. Out of a total of 68 samples tested, 29% reacted positively with BYDV-PAV antiserum, 24% with BYDV-RPV antiserum and 7.4%, with both PAV and RPV antisera (Table 2). The remaining 39.6% reacted with neither antiserum. The BYDV-PAV antiserum did not react with BYDV-RPV (Table 2). The colour intensities that developed in some of the positive sample wells were similar to those in positive control wells (Table 2). No colour development was observed in wells that contained sap from healthy plants. Disease incidence. Since it was not possible to distinguish the BYDV-PAV and RPV serotypes based on symptoms, the proportion of infected plants was that showing BYDV symptoms. The overall average disease incidence in the surveyed irrigated wheat fields in the Central, Lusaka and Southern provinces ranged between 5% and 25% with the highest infection level (25%) recorded in Central and Lusaka provinces. Distribution of BYDV serotypes PAV and RPV in the wheat fields. BYDV serotypes PAV and RPV were recovered from 9 and 10 of the 11 surveyed fields, respectively. The proportion of infections with BYDV serotypes PAV and RPV varied considerably in the wheat fields (Fig. 1). The highest number of samples containing serotype PAV was recorded in wheat field No. 4 (G. Bender), whereas samples collected from fields Nos. 1 (Chindindindi) and 8 (Muyiuku) did not contain BYDV serotype PAV (Fig. 1). BYDV serotype RPV was found in a relatively large number of samples from field 4, while in field 11 (Pestol), none of the samples tested contained this serotype (Fig. 1). Of the 9 wheat cultivars surveyed 4 were infected with serotype PAV, 1 with RPV and 4 with both. DISCUSSION The BYDV complex is important on all the small-grain cereals (Brunt et al., 1990), but information on its occurrence in Africa is very scanty (Thottappilly, 1992). In this study, results showed that both BYDV PAV and RPV infect wheat in Zambia. PAV was the more commonly detected serotype (Table 2). In Morocco and Tunisia, the PAV type of BYDV was also more commonly detected (Makkouk et al., 1992; Yaman, 1992) in wheat population as in most cereal growing areas of the world (Plumb, 1974; Rochow et al., 1986). The fact that some symptomatic leaf samples gave no reaction to either antiserum suggests that they were infected with virus(es) different from BYDV-RPV and -PAV. Symptoms induced by both serotypes were indistinguishable in the fields. The stunting of infected plants and leaf chlorosis observed in this study correspond to previous findings (Wiese, 1987; Brunt et al., 1990). BYDV cause leaf distortion, curling and shortening of stem internodes and poor heading in the infected wheat (Bruehl, 1961) as noticed in the present study. Burnett and Gill (1976) have reported the progressive shortening of main culm and heads of BYDV-infected wheat plants. Such symptoms were observed in some wheat cultivars during this study. Because luteoviruses are phloem-limited, mechanical transmission to indicator hosts and serological techniques that are less sensitive than ELISA are not useful diagnostic tools (D’Arcy et al., 1989). In the present study, DAS ELISA proved to be useful for studying BYDV. The antisera used in the present study could effectively distinguish BYDV serotypes PAV and RPV, and both BYDV serotypes were identified occasionally co-infecting some wheat plants in surveyed fields. Burnett and Mezzalama (1992) also detected mixed infection of BYDV serotypes PAV and RPV in a wheat population using ELISA. Rochow (1979b) reported that mixed infection of BYDV serotypes caused a synergistic increase in symptom severity (Rochow, 1970b). Very severe symptoms were also observed in plants co-infected with BYDV serotypes PAV and RPV in this study (Table 2), suggesting the synergistic effect of the two serotypes. Casper (1988) reported that five BYDV serotypes infect wheat, namely MAV, PAV, SGV and RMV. In the present study however, only PAV and RPV could be investigated due to limited funds to purchase MAV, SGV and RMV specific antisera. The average barley yellow dwarf disease incidence in irrigated wheat fields in the three provinces ranged from low to high (5% to 25%). Makkouk et al. (1992) also reported BYDV incidence ranging between 2% to 30%, and Burnett and Mezzalama (1992) of up to 35%, in infected wheat which was considered to be high. The virus serotypes PAV and RPV were evenly distributed in the fields in the present study (Fig. 1). With the continuous growing of the same wheat cultivars in these areas, and depending on the status of BYDV serotype vectors, the virus might spread even more and cause a serious disease outbreak that could result in major yields and quality losses in wheat grown in Zambia. CONCLUSIONS In this study, BYDV serotypes PAV and RPV were found infecting irrigated wheat in Zambia for the first time. In crop disease management programmes, efforts must be made not only to identify the pathogens but also to recognise important variation (serotypes) in their populations as mild and severe virus variants/serotypes are constantly evolving and their detection provide information on which to base control measures. The possibility that other BYDV serotypes (MAV, RMV and SGV) exist in irrigated wheat in Zambia cannot be ruled out, and need to be studied for the symptomatic leaf samples which reacted negatively to PAV and RPV antiserum in this study could contain these serotypes. The role of alternative hosts and host-virus vector interactions in the epidemiology of BYDV serotypes should also be investigated, since little is known about the host range and the epidemiology of BYDV in the tropics (Thresh, 1998). Furthermore, response of host plants to the BYDV serotypes need to be evaluated so that natural resistance can be identified and exploited and novel resistance mechanisms developed to halt the spread of BYDV in Zambia. ACKNOWLEDGEMENTS We thank CIMMYT, Harare, Zimbabwe for the financial support and Ms. Olga Kamanga, Tissue Culture Laboratory, University of Zambia for her technical and material support. We also thank Dr. Richard Gibson of NRI, for his comments on the manuscript. REFERENCES
The following images related to this document are available:Line drawing images[cs00020a.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}