 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 8. No. 4, pp. 355-364 Ploidy level of the cultivated yams (Dioscorea cayenensis/D. rotundata complex) from Benin Republic as determined by chromosome counting and flow cytometry A. Dansi1,2, M. Pillay3, H.D. Mignouna3, O. Daïnou1, F. Mondeil4 and K. Moutaïrou5 (Received 17 July, 2000; accepted 20 October, 2000) Code Number: CS00038 INTRODUCTION Tropical root and tuber crops occupy a pre-eminent position as food crops, next only to cereals and grain legumes, and they also form the subsidiary staple of over 20% of world population (Okwor, 1998). Among the tropical tuber crops, Guinea yam (Dioscorea cayenensis/Dioscorea rotundata complex) is one of the most important, especially in the so-called ‘yam belt’ of West Africa (Orkwor et al., 1998). In spite of its economic importance, Guinea yam has not received the attention that it deserves with regard to its quantitative and qualitative improvement. Consequently, many genotypes are reported to be susceptible to pests and diseases (Degras, 1986; Dansi et al., 1997, Orkwor et al., 1998). For developing new elite genotypes for ecological adaptation and resistance to pests and diseases, plant breeders will need access to a wide range of diversity. Therefore, better knowledge of the existing traditional cultivars held by farmers is a pre-requisite. In order to assess the diversity within this species complex (D. cayenensis /D. rotundata) in the Republic of Benin, systems of classification and identification based on morphological, isozymic and RAPD markers were recently used (Dansi et al., 1998, 1999, 2000a, b). D. cayenensis /D. rotundata complex being polyploid, the knowledge of the ploidy level of the cultivars identified within this germplasm is important. Determining ploidy levels in yam by counting chromosomes is tedious, difficult and time consuming. Yam chromosomes are small, generally dot-like and most often clumped together complicating the counting (Baquar 1980; Zoundjihékpon et al., 1990). To overcome these difficulties, flow cytometry has been recently used to determine ploidy levels in yams (Hamon et al.,1992; Gamiette et al., 1999). In ploidy analysis, flow cytometry assay has some important advantages over conventional chromosomes counting. The method is non-destructive (one sample can be prepared from a few milligrams of leaf tissue), exceptionally rapid, sensitive and convenient, does not require dividing cells, and can detect both mixoploidy and aneuploidy (Galbraith et al., 1983; De Laat et al., 1987; Arumuganathan and Earle, 1991a, b; McMurply and Rayburn, 1991; Dolezel, 1997). The objective of this study was to determine the ploidy level of the different cultivars identified within the D. cayenensis /D. rotundata complex of Benin Republic using both chromosome counting and flow cytometry. MATERIAL AND METHODS Plant materials. The material consisted of 90 yam cultivars of the D. cayenensis/D. rotundata complex of Benin Republic (Tables 1 and 2). These cultivars represent the ninety morphotypes identified within the Guinea yam germplasm collected in 1996 and 1997 in different localities of Benin (Dansi et al., 1997; 1998; 1999). They are maintained as a field collection at the International Institute of Tropical Agriculture near Cotonou.
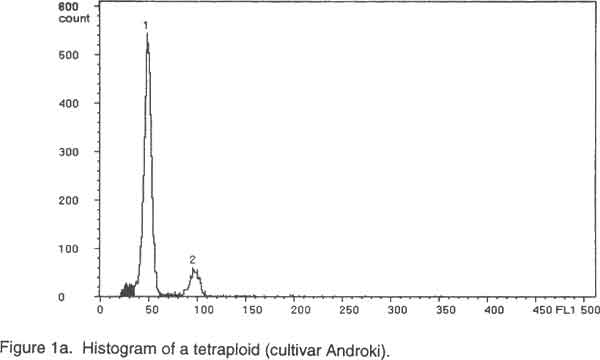
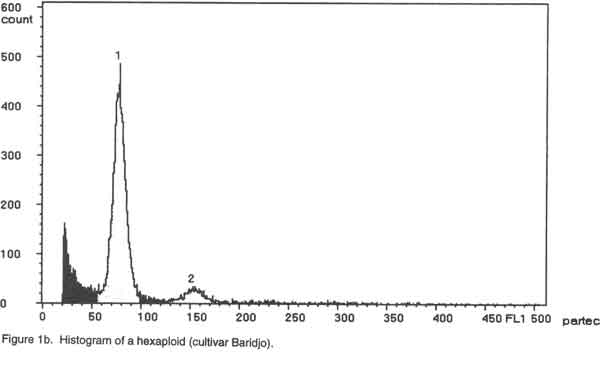
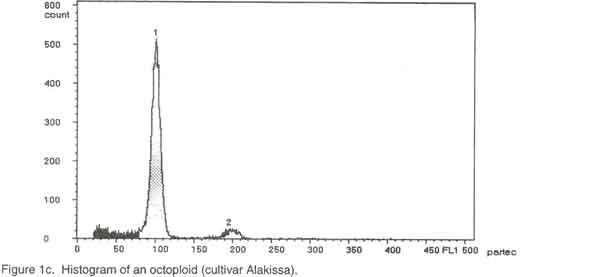
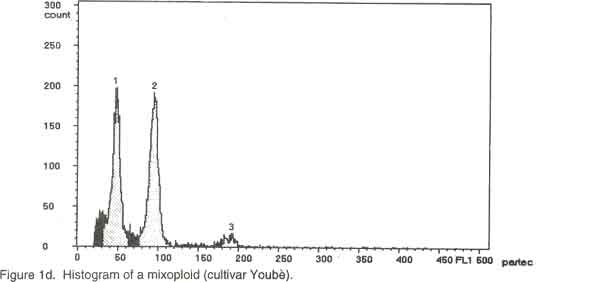
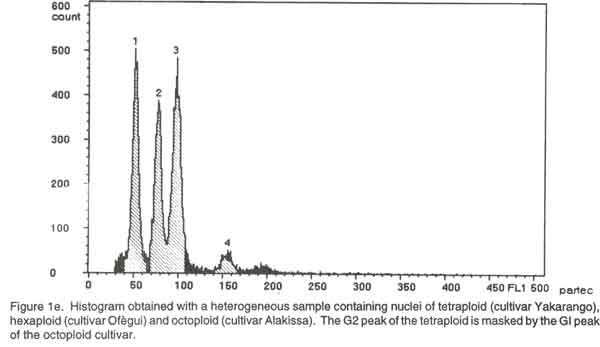
Of the ninety cultivars selected, 26 were analysed by chromosome counting (Table 1), 60 by flow cytometry (Table 2) and 4 by both chromosome counting and flow cytometry (Table 2). The 30 cultivars analysed by chromosome counting (Table1) were selected in such a way that the 26 cultivar groups identified within the Benin Republic’ Guinea yam germplasm were represented. For the purpose of the analysis, plants were cultured in vitro (nodal cutting culture) on the basal MS medium (Murashige and Skoog, 1962), supplemented with 2% sucrose, 0.05 mg/l kinetin, 20 mg/l cysteine and 0.7% agar, pH= 5.6 at 25°/ 22°C day/night. The media were sterilised at 121°C for 15 minutes and cooled before use. Lighting was provided by cool white fluorescent lamps (18 h photoperiod and 4000 lux light intensity). Chromosome counting. Slides were prepared by the technique outlined by Zoundjihékpon et al., (1990). The root tips were obtained soon after sunrise (before 7 a. m.) from plants grown in sawdust in a greenhouse. Roots were pre-treated in a 2mM 8-hydroxyquinoline solution for about 6 h on filter paper in a petri dish to accumulate metaphase cells. The roots were fixed in acetic –alcohol (1/3) for 48 h, followed by hydrolysis in 5N HCl for 45 min and 1N HCl for 10 min. Root tips were stained in Feulgen reagent in darkness for at least 2 h and slides were prepared by squashing in 45 % acetic acid. Prepared slides were stored at -20°C. After removing the cover slip, the slides were soaked in absolute ethanol for 1 h, air-dried for 15 min, stained in a Giemsa solution in phosphate buffer, pH=6.7, rinsed in distilled water for 2 min and dried again before mounting. The chromosomes were counted in 3-5 cells per slide and in 5-10 root tips per cultivar using a light microscope at magnification of 1000x. Preparation of nuclei for flow cytometry. A procedure modified from Otto et al. (1981) and Otto (1990) was used to prepare and stain the cell nuclei. To release nuclei, young and healthy plant leaves were cut into pieces in a glass petri dish. Following addition of 1 ml of ice-cold extraction buffer (0.1M citric acid containing 0.5% Tween 20), the leaf pieces were finely chopped with a sharp razor blade. The homogenate was filtered through a 45 µm nylon gauze filter to remove cell debris. Two ml of staining buffer containing 0.4 M sodium hydrogen phosphate and 4 µg/ml DAPI (4'-6-diamidino-2-phenylindole) was added to the suspension of nuclei and samples were analysed immediately. Flow cytometry. Flow cytometry measurements were performed with Partec Ploidy Analyser (Partec Gmbh, Germany) equipped with a 100W high-pressure mercury lamp. Instrument gain was adjusted so that the G1 peak of nuclei isolated from a control tetraploid plant (cultivar Kponan with a chromosome number of 40) was set at channel 50. This calibration was checked periodically to minimise variation due to runs and kept constant during the analysis of samples prepared from plants of unknown ploidy. Peaks representing G1 nuclei were then expected at channels 75 and 100 for hexaploid and octoploid ploidy levels respectively. Two measurements were made for each isolation and at least 2000 nuclei were examined each time. To estimate ploidy level, the position of the G1 peak on the histogram obtained for each of the individuals was compared to that of the cultivar ‘Kponan’. A software package (Partec) was used for the calculation of CV-values. RESULTS Chromosome counts. Among the thirty cultivars investigated, two (Baridjo and Makpawa) were hexaploid and showed 60 chromosomes, one (Agangan) was octoploid with 80 chromosomes and the others were tetraploid with 40 chromosomes (Table 1). In cultivar ‘Makpawa’, some cells showed 62 and 63 chromosomes within the same root tips while counts of about 65 were observed infrequently. One or two extra chromosomes were also observed in some cells of the tetraploid cultivars ‘Antawororou’, ‘Djikpiri’, ‘Guiena’ and ‘Issou agatou’. In all the cultivars analysed, chromosomes were small with some appearing dot-like and others rod-shaped without any visible centromere region. In most of the cells, chromosomes appeared clumped together hence complicating counting. Flow cytometry. After DAPI staining, isolated nuclei irradiated with UV radiation from young leaf tissue emitted fluorescence which was measured by the flow cytometer. For each sample, analysis of the relative fluorescence intensity of the isolated nuclei yielded a histogram showing a dominant peak corresponding to nuclei in the G1 phase of the cell cycle and a minor peak corresponding to G2 nuclei. The amount of debris in the samples was negligible. The dominant G1 peaks made ploidy estimation easy. Three ploidy levels, tetraploid (4X), hexaploid (6X) and octoploid (8X) were observed among the sixty-four cultivars analysed (Table 2). Fifty-three cultivars were tetraploid (Fig. 1a), five were hexaploid (Fig. 1b) and three were octoploid (Fig.1c). Two cultivars Youbè and Tam-Sam were mixoploid (Fig. 1d). The relative nuclear DNA content in arbitrary units (AU), expressed as channel numbers varied from 43,7 AU to 52,2 AU for the tetraploids, from 65,4 AU to 76,9 AU for the hexaploids and from 85,5 AU to 100,3 AU for the octoploids (Table 2). The coefficient of variation (CV), determined as the quotient of standard deviation of the peak and the mean peak position (channel number), was 3.4 to 4.7% (mean = 3.8) for the G1 peaks of plant nuclei (Table 2). The small CVs reflected narrowness of the peaks and indicated good reliability of measurements. Clear separation of peaks was even obtained when a sample was prepared from a mixture of leaves from tetraploid, hexaploid and octoploid individuals (Fig. 1e). DISCUSSION Dioscorea is one of the most difficult genera for cytotaxonomic and cytogenetic studies (Essad, 1984). Many authors already reported the difficulties encountered in chromosome counting in cultivars of D. cayenensis/D. rotundata complex (Miège 1952, 1954; Baquar, 1980; Zoundjihékpon et al., 1990) and even other Dioscorea species (Suessenguth, 1921; Ramachandran, 1968; Baquar, 1980; Araki et al., 1983; Essad, 1984, Gamiette et al., 1999). In yam, the presence of extra chromosomes in the cells of some individuals as observed in this study is not rare and has been already reported by Miège (1954), Baquar (1980), Zoundjihékpon et al., (1990) and Gamiette et al. (1999). The extra chromosomes are reported to be B chromosomes or satellites which in yam are sometimes as large as the chromosomes themselves (Essad, 1984). Off the 90 cultivars analysed (Tables 1 and 2), eighty were tetraploid, three were octoploid, five were hexaploid and two were mixoploid. When considering the cultivar groups, twenty-three out of the twenty-six investigated appeared tetraploid while one (Alakissa) was octoploid and two (Baridjo and Makpawa) were hexaploid. As found in Côte d’Ivoire (Zoundjihékpon et al., 1990; Hamon et al., 1992) and Cameroon (Dansi et al., 2000c), the tetraploids form the largest group in Benin Republic (Fig. 2). This is in agreement with the findings of Essad (1984) which indicated that tetraploid individuals are the most frequent in the Dioscorea species. The diverse chromosome counts reported by Miège (1954), Sharma and De (1956), Martin and Ortiz (1963), Baquar (1980) and Essad (1984) in the D. cayenensis /D. rotundata complex, indicated the existence of two chromosome base numbers, X= 9 and X=10. Only the chromosome base number X=10 was observed in the Benin Guinea yam cultivars. Similar results have already been reported for yams in Côte d’Ivoire and Cameroon (Zoundjihékpon et al., 1990). DAPI is an AT base pair binding fluorescent dye, and is not suitable for DNA content estimation in plants (Michaelson et al., 1991; Dolezel et al., 1992; Godelle et al., 1993; Dolezel et al., 1998). The Partec ploidy analyser used in this study only had filters for DAPI, so it was not possible to analyse the nuclear DNA content variation (in absolute units) within the yams of Benin. However, variations in the relative fluorescence intensity (Channel number) recorded in each of the classes of ploidy level may be an indication of a probable variation in the nuclear DNA content (in absolute unit) in the samples. Such intraspecific variation of nuclear DNA content are frequent in plants and have been already reported in many plants such as maize (Laurie and Bennett, 1985; Rayburn et al., 1989; Biradar and Rayburn, 1993), rice (Martinez et al., 1994), and soybean (Hammatt et al., 1991). Analysis of the nuclear DNA content variation in the Benin yams may be important because it could be correlated with the flowering capacity and some agronomic traits of the plants as shown in potato by Valkonen et al. (1994). In this study, no correlation was found between ploidy level and geographic origin of the yam cultivars in the Republic of Benin. This is contrary to the study of Miège (1954) who described local geographic centres of origin for the tetraploids, the hexaploids and the octoploids in Côte d’Ivoire. Some authors (Chevalier, 1936, 1946; Burkill, 1960; Miège, 1968) treat D. cayenensis and D. rotundata as distinct species while others (Hamon, 1987; Ramser et al., 1997; Dansi et al., 1999, 2000a) consider the two as different compartments of the same species. In the present study, a correlation was found between subspecies or compartments and ploidy levels. Hence, all the cultivars (Agangan’, ‘Alakissa’, Baridjo’, Doundoua’, ‘Makpawa’, Ofegui’, ‘Ouwon-peotina’, ‘Sogodo’) belonging to the subspecies D. cayenensis are either hexaploids or octoploids while those of D. rotundata are all tetraploids. Because the plant material used in the flow cytometry analysis were cultured in vitro, chromosome counts are needed to confirm the mixoploid status of the two cultivars ‘Youbè’ and ‘Tam-Sam’ in which two populations (4X and 8X) of cells were detected. In fact, such results may be the consequence of endomitosis that has occurred in some cells during the growth in vitro. The presence of 40 chromosomes, in all the cells investigated in the cultivar ‘Iberegnense’ that is morphologically identical to ‘Tam-Sam’ supports the above point of view. This investigation is the first report of cytogenetic work on the cultivated yams belonging to D. cayenensis /D. rotundata of Benin Republic. It is an important prebreeding examination of the cultivars which will direct the yam breeding programme both towards the choice of initial material and towards the breeding methods that will supply the modern cultivars. CONCLUSION Guinea yams (D. cayenensis/D. rotundata complex) in the Republic of Benin, like in Côte d’Ivoire, Nigeria and Cameroon, are also polyploid with tetraploid, hexaploid and octoploid individuals. Among these, tetraploids individuals are the most frequent. In yam breeding programme, an accurate knowledge of the ploidy level of the cultivars is required. Because of the difficulties in chromosome counting previously highlighted, the use of flow cytometry which gave result in agreement of chromosome counts offers the most suitable tool for this purpose. The ninety different cultivars for which ploidy levels were determined will be evaluated, multiplied and used for either breeding or other genetic investigations. This will help create new elite genotypes for improved yam productivity in Benin and other parts of West Africa. ACKNOWLEDGEMENTS We thank the International Foundation for Science (IFS) for financial support. The technical assistance of Tunday Adeoshun and Monday Odishiwo (IITA, Ibadan) throughout the study is acknowledged. We are also grateful to all the farmers we met for sharing their material and fruitful discussions while collecting and classifying the yams. REFERENCES
The following images related to this document are available:Photo images[cs00038d.jpg] [cs00038a.jpg] [cs00038e.jpg] [cs00038b.jpg] [cs00038c.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}