 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
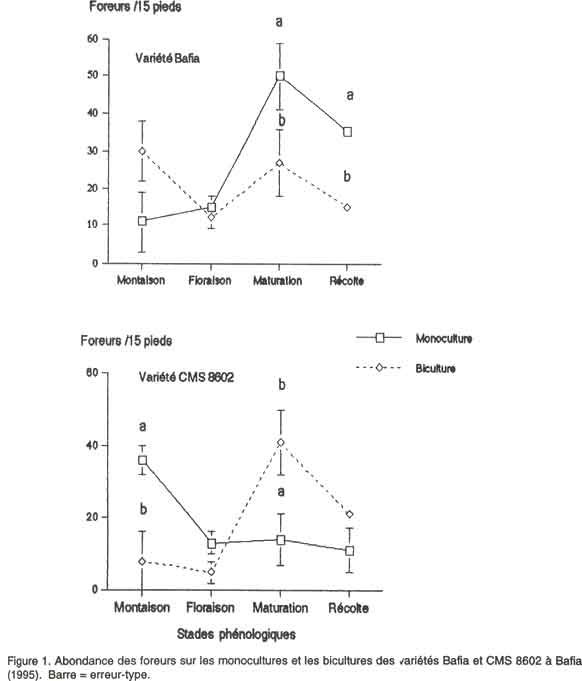
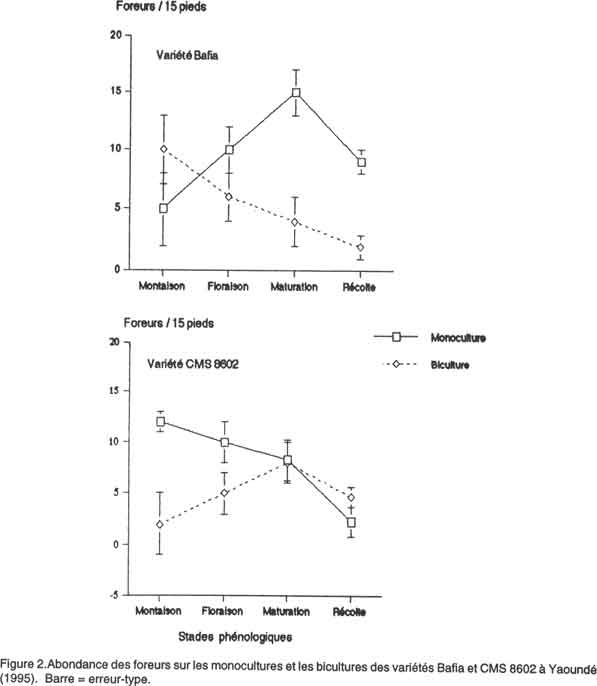
African Crop Science Journal, Vol. 8. No. 4, pp. 365-374 Abondance et diversité des foreurs de tiges et grains dans une biculture maïs-arachide au centre du Cameroun A. Régine et D. Coderre1 (Received 3 May, 1999; accepted 3 July, 2000) Code Number: CS00039 INTRODUCTION Les attaques des lépidoptères foreurs des tiges et graines constituent l'une des principales contraintes à la culture du maïs au Cameroun. Les principales espèces sont: les noctuelles Busseola fusca (Fuller) et Sesamia calamistis (Hmps), ainsi que les pyrales Eldana saccharina (Walker) et Mussidia nigrivenella (Ragonot) (Aroga, 1987a, 1987b; Cardwell et al., 1993). La majorité des larves vivent à l'intérieur des tiges ou des épis (Aroga, 1987b; Conlong et al., 1994). Ce comportement, qui limitant ainsi l'efficacité de la lutte chimique justifie probablement les augmentations des quantités et les fréquences d'application des insecticides (Seshu Reddy, 1983; Kfir, 1990). Cependant, les effets néfastes de la lutte chimique sur les humains et sur l'environnement, ainsi que leur coût de plus en plus élevé, accentuent l'importance d'envisager la réduction de leur usage à un niveau économiquement et environnementalement acceptable par le biais de l'application des stratégies alternatives, notamment les pratiques culturales, la lutte biologique, la résistance variétale, etc. La polyculture est une pratique culturale courante de l'agriculture camerounaise. Dans le centre, le maïs est souvent associé à l'arachide (Arachis hypogaea), qui est une des principales sources de protéines végétales (les graines ont une teneur de 25% en protéines) des populations locales (Gillier et Silvestre, 1969; Leplaideur, 1985; Ay et al., 1986). Des travaux expérimentaux en station vérifiant l'influence de cette pratique ont révélé que des augmentations de rendement de maïs de l'ordre de 11 à 21% pouvaient être obtenues dans les parcelles de maïs associé aux arachides par rapport aux monocultures de maïs correspondantes (Mutsaers, 1976). Cependant, très peu d'efforts ont été orientés vers l'examen de la réponse d'insectes ravageurs à ce système cultural. Selon les travaux de plusieurs chercheurs, notamment Root (1973), Risch (1979) et Trenbath (1993), la polyculture réduit le nombre de certains herbivores et renforce l'abondance de leurs ennemis naturels. L'hypothèse de "la concentration de ressources" proposée par Root (1973) prévoit une réduction de l'aptitude des herbivores (surtout les spécialistes comme les pucerons ou les doryphores) à découvrir et utiliser efficacement leurs hôtes dans les milieux de cultures diversifiées. La polyculture pourrait affecter l'usage des stimuli visuels par lesquels les insectes reconnaissent leurs plantes-hôtes. Waterworth (1994) a rapporté qu'il y avait moins d'attaque sur le coton associé à l'arachide dans l'Est de la Zambie. Coderre et al. (1989) ont enregistré un plus petit nombre de pucerons Metopolophium dirhodum (Walker) dans les bicultures maïs-haricot. La polyculture peut aussi perturber les stimuli olfactifs. Ainsi, Podagrica sp. a été repoussé sur le coton et Plutella xylostella L. sur le chou par les odeurs des plantes aromatiques telles que l'aïl, l'oignon ou la tomate associées à leurs hôtes (Tahvanainen et Root, 1972). Les taux d'immigration et d'émigration des herbivores peuvent aussi diminuer dans les systèmes diversifiés. Elmstom et al. (1988) ont signalé un important taux d'immigration de Phyllotreta cruciferae Goeze dans les monocultures de brocoli par rapport aux bicultures brocoli-trèfle blanc. Litsinger et al. (1991) ont signalé que les plantes non hôtes associées au maïs constituaient une barrière efficace à la dispersion des jeunes larves du foreur Ostrinia furnacalis (Guenée). L'objectif de la présente étude était d'évaluer l'abondance, la diversité et l'activité des populations de foreurs dans une biculture maïs-arachide au centre du Cameroun. L'étude a été conduite à la fois dans la partie forestière et dans la zone de savane. Des monocultures et bicultures maïs-arachide de deux variétés de maïs (une locale et une améliorée) étaient comparées. MATÉRIEL ET MÉTHODES Des parcelles agricoles ont été installées à Yaoundé (3°52' latitude N et 11°26' longitude E) et à Bafia (4°38' latitude N et 11°23' longitude E). Deux variétés de maïs à cycle court (90 jours en moyenne), le CMS 8602 ("Cameroon Maize Selection", deuxième campagne, 1986) et le Bafia (une variété locale), étaient comparées Chacune des variétés était semée sur des parcelles de monoculture et de biculture de 10 m x 10 m. On obtenait ainsi quatre traitements : le Bafia en monoculture (LM) et en biculture (LB), puis le CMS 8602 en monoculture ( AM) et en biculture (AB). Le maïs des monocultures et des bicultures était semé à des écartements de 0,25 m sur la ligne et 0,80 m entre les lignes (50 000 pieds ha-1). Les semences qui n'avaient pas été préalablement traitées, étaient implantées sur des blocs séparés les uns des autres par des allées de 2 m de large. Dans les bicultures, trois lignes d'arachide étaient intercalées entre deux lignes de maïs. Les pieds d'arachide se trouvaient à 0,30 m les uns des autres entre les lignes, et à 0,10 m sur les lignes (333.000 plants ha-1). Le plan expérimental était un bloc aléatoire complet en trois répétitions. Aucun insecticide n'a été appliqué. Cependant, un engrais composé NPK (20-10-10) à la dose de 600 kg ha-1 et un herbicide de pré-levée, le gramoxone Super (800g m. a. ha-1), ont été appliqués au semis. Un remplacement des plants manquants a été effectué à la levée (une semaine après le semis initial). Un désherbage manuel a été effectué sept semaines plus tard. Un échantillonnage bihebdomadaire de trois pieds de maïs arrachés ou sectionnés au niveau du collet au hasard sur chacune des six lignes centrales était effectué; ces plantes étaient ensuite disséquées, et le nombre d'oeufs, de larves et de chrysalides de foreurs présents noté. La proportion des coeurs morts et le degré de dégâts sur les feuilles ont été évalués à la montaison sur quinze pieds de maïs choisis au hasard dans chaque parcelle. La sévérité des attaques sur les feuilles était évaluée à l'aide de l'échelle de Guthrie (Kumar, 1993) graduée de 1 (aucun trou visible) à 9 (plupart de feuilles avec de longues lésions). À la récolte, la longueur des galeries de quinze autres pieds choisis au hasard dans les six lignes centrales a été mesurée, afin de calculer la proportion de la tige creusée. Le poids des épis des trois lignes centrales a été relevé, et le rendement en grains secs calculé selon la formule Y = FW x SP x (100 - FMC) / 85 x 1 HA / LA, où FW = le poids des épis au champ immédiatement après la récolte ; SP = pourcentage d'égrenage assimilé à 0,83 ; FMC = le taux d'humidité des grains à la récolte (%) ; 100 - FMC = le poids de la matière sèche par ligne de récolte (kg) ; 1HA = la superficie d'un hectare (m2) ; LA = la superficie occupée par les lignes récoltées ; 85 = la constante dérivant du taux de référence des grains à la récolte (15%) reconnu par le CIMMYT. Afin d'évaluer l'impact de la biculture sur la migration des larves de foreurs, cinq larves néonates de B. fusca/pied ont été placées sur deux pieds centraux de chacune des dix micro-parcelles (cinq de monocultures et cinq de bicultures) de 1m x 1m, installées à cet effet. La distance parcourue par ces larves a été suivie pendant 1h 30 min. Analyses statistiques. Les différentes données collectées ont été soumises à une analyse de variance à trois critères de classification (zone écologique, variété de maïs et système cultural) afin de déceler les effets de chacun des facteurs. L'indice de diversité de Shannon-Weiner, calculé à partir de l'abondance absolue des espèces de foreurs selon la formule H' = -∑ (pi)(log pi) (où pi = nombre d'individus d'une espèce / nombre total d'individus) (Peet, 1974), et l'indice de régularité de leurs distributions de fréquence ("evenness"), mesuré par la formule J' = H' / logS (où S = nombre d'espèces dans la communauté) (Krebs, 1989), ont servi à la comparaison de leur diversité dans les différents systèmes. Les pourcentages de coeurs morts et de galeries dans la tige ont été corrigés à l'aide de la racine carrée d’arc sinus avant l'analyse. Le degré de dégâts sur les feuilles a été soumis au test non paramétrique de Kruskall-Wallis. RÉSULTATS Importance relative et diversité des espèces de foreurs. B. fusca, S. calamistis, E. saccharina et M. nigrivenella ont été les espèces de foreurs recensées sur le maïs lors de notre étude (Tableau 1). Toutes étaient présentes à Yaoundé, alors que E. saccharina était absente à Bafia, et la proportion de M. nigrivenella y était insignifiante. Ces espèces ont été collectées sur les deux variétés testées. Cette même composition spécifique a été notée dans les monocultures et dans les bicultures. Les indices de diversité spécifique et la régularité de distribution de fréquence relevés à Yaoundé étaient plus importants que ceux observés à Bafia (Tableau 1). Ils étaient presqu'identiques sur les deux variétés de maïs, sauf sur les monocultures à Yaoundé où un indice plus grand a été relevé sur le CMS 8602. Les indices des monocultures étaient généralement plus petits que ceux des bicultures.
L'abondance relative de B. fusca était significativement plus grande à Bafia qu'à Yaoundé (P < 0,05) (Tableau 1). Celle de E. saccharina l'était plutôt à Yaoundé (P < 0,001). Il n’y avait pas d'écart significatif entre les abondances relatives de S. calamistis et de M. nigrivenella des deux localités. Il n'y avait pas de différence significative entre les abondances des espèces sur les deux variétés et dans les deux systèmes culturaux (Tableau 1). B. fusca était l'espèce prédominante dans les deux localités, sur les deux variétés et sur les deux systèmes de culture. Elle constituait avec S. calamistis, les seules composantes de la population de foreurs jusqu'au stade de floraison. E. saccharina et M. nigrivenella s'y sont ajoutées après cette époque. Abondance des foreurs. Le nombre de foreurs sur la variété Bafia était significativement (P<0,0001) plus grand à Bafia qu'à Yaoundé, quelque soit le système cultural considéré. Les monocultures des deux localités en portaient moins que les bicultures à la montaison, mais les différences n'étaient pas significatives (Fig. 1 et Fig. 2). Après la floraison, l'abondance des foreurs sur les monocultures était supérieure à celle des bicultures. Toutefois, des différences significatives (P< 0,01) n'ont été notées qu'à Bafia à la maturation et à la récolte (P< 0,05) (Fig. 2). Sur la variété CMS 8602, le nombre de foreurs était significativement (P<0,001) plus grand à Bafia qu'à Yaoundé, quelque soit le système cultural. À la montaison, ils étaient plus abondants sur les monocultures que sur les bicultures des deux localités, avec une différence significative uniquement à Bafia (P< 0,05) (Fig. 2). À la maturation par contre, le nombre de foreurs était significativement (P< 0,05) plus grand sur les bicultures à Bafia (Fig. 2). Migration des larves des foreurs. Six pour cent des 50 larves néonates de B. fusca introduites dans les micro-parcelles de bicultures maïs-arachide ont réussi à migrer sur les pieds voisins 1 h30 min après leur introduction. Un pour cent seulement du même nombre, introduit dans les micro-parcelles de monocultures a eu un comportement semblable pendant cette même durée. Toutes les larves migratrices des bicultures se sont retrouvées sur les pieds d'arachide immédiatement voisins à ceux de maïs, et aucune n'est pas allée au-delà. Les migratrices des monocultures ont rejoint les pieds de maïs voisins. Alors que la mortalité de celles-ci pendant ce laps de temps était nulle dans les monocultures, elle atteignait un taux de 100% dans les bicultures. Dégâts des foreurs et rendement en grains secs. La proportion des coeurs morts était plus faible à Yaoundé qu'à Bafia, toutefois, l'écart n'était pas significatif entre les deux localités (Tableau 2). Sa valeur était plus forte sur la variété locale à Bafia par rapport au CMS 8602, mais la différence était non significative. La proportion des coeurs morts des bicultures était généralement supérieure à celle des monocultures, avec des écarts non significatifs .
Les attaques sur les feuilles étaient plus sévères à Bafia qu'à Yaoundé, avec une différence non significative (Tableau 2). Cette sévérité n'a pas été influencée de façon significative ni par la variété ni par le système cultural. La proportion de la tige occupée par une galerie était significativement (P = 0,0001) plus importante à Bafia qu'à Yaoundé (Tableau 2). Il n'y avait pas d'écart significatif entre ceux relevés sur les deux cultivars, et la différence entre celle des monocultures et celle des bicultures n'était pas importante. Le rendement en grains secs a été significativement (P = 0,0001) meilleur à Yaoundé qu'à Bafia (Tableau 2). Des différences non significatives étaient notées entre les variétés. Il n'y avait pas d'écart important entre les systèmes culturaux, sauf sur la variété Bafia à Yaoundé où un meilleur rendement a été noté sur les bicultures. DISCUSSION La présence du complexe de quatre espèces de foreurs de maïs B. fusca, S. calamistis, E. saccharina et M. nigrivenella a déjà été signalée au Cameroun et particulièrement dans la zone étudiée (Cardwell et al., 1993). Ce complexe était présent à Yaoundé sur les deux variétés de maïs et sur les deux systèmes culturaux (Tableau 1). À Bafia par contre, E. saccharina était absente dans tous les systèmes. Les indices de diversité et de régularité de ces espèces sur les monocultures et sur les bicultures d'une part, sur le CMS 8602 et sur le Bafia de l'autre à Yaoundé étaient supérieurs à ceux des systèmes correspondants à Bafia, indiquant une répartition plus équitable de celles-ci dans la première localité . Selon Legendre et Legendre (1979), des faibles indices de régularité indiqueraient une activité biologique plus élevée. Ainsi, les faibles indices enregistrés à Bafia semblent indiquer une préférence de certaines espèces pour cette zone écologique. Effectivement dans cette dernière, une seule espèce représentait plus de 95 % de la population totale (Tableau 1). On pourrait ainsi conclure que l'écosystème forestier a été plus favorable à la diversité des espèces de foreurs que celui de savane. Des indices de diversité spécifique presqu'identiques sur les deux cultivars indiquent que la variété n'a pas influencé la répartition. Des indices plus forts relevés sur les bicultures par rapport aux monocultures suggèrent une meilleure répartition dans les premières que dans les secondes. B. fusca était l'espèce prédominante du complexe, et ce, quelque soit la localité, la variété ou le système cultural. Ceci appuie les résultats des études faites dans d'autres régions. Effectivement, il est considéré comme le foreur le plus important sur le maïs et le sorgho dans la plupart des régions d'Afrique au sud du Sahara et en Éthiopie (Tchekmenev, 1981; Seshu Reddy, 1983). Les époques d'apparition de ces espèces indiquées dans nos résultats, correspondent à celles rapportées ailleurs (Seshu Reddy, 1983; Aroga, 1987a; Bosque-Pérez et Mareck, 1990). Elles n'ont pas été modifiées, ni par la zone écologique, ni par la variété, encore moins par le système cultural. La variation temporelle du nombre de foreurs présente une courbe avec deux maxima. Le premier qui apparaît au stade de montaison, correspond à une accumulation des larves de première génération de B. fusca et de S. calamistis. Le second et le plus important, intervient au stade de maturation, et coïncide avec l'époque d'apparition de E. saccharina et M. nigrivenella. L'importance de ce pic est renforcée par l'arrivée des larves de deuxième génération de B. fusca et de S. calamistis parmi les populations. Le creux entre les deux sommets serait la conséquence d'une forte mortalité intervenue autour de l'époque de floraison, lorsque les larves forcées à quitter le cornet, se sont exposées à la dessiccation et aux ennemis naturels (Appert et Deuse, 1982 ; Aroga, 1987b ; Berger, 1992). Selon certains auteurs, la polyculture masque les stimuli visuels et / ou olfactifs par lesquels les insectes reconnaissent leurs plantes-hôtes. Par exemple, Tahvanainen et Root (1972) ont rapporté que l'odeur de la tomate associée au coton ou au chou a repoussé Podagrica sp. sur la première plante et P. xylostella sur la seconde. Les foreurs insectes actifs surtout la nuit, sont attirés par les odeurs émises par leurs plantes-hôtes (Renwick et Radke, 1983; Schneider, 1987). Le nombre de larves de foreurs significativement plus élevé dans les monocultures (surtout à la montaison et à la maturation) (Figs. 1 et 2) suggère que la majorité d'oeufs ont pu être déposés dans ces systèmes. On pourrait donc supposer que la présence de l'arachide a perturbé les femelles pondeuses lors de leur quête des lieux favorables pour la ponte. Dans ce cas, les volatiles émis par l'arachide auraient masqué ceux du maïs dans les bicultures, contribuant de ce fait à repousser les pondeuses. D'où le plus faible nombre de larves relevé dans ces systèmes. Selon Kareiva (1985), la concentration de ressources réduit le taux d'immigration et d'émigration des herbivores. Litsinger et al. (1991) ont signalé que les plantes non hôtes associées au maïs constituaient une barrière efficace à la dispersion des jeunes larves du foreur O. furnacalis. Dans nos micro-parcelles de biculture, les larves de B. fusca réussissaient rarement à traverser les pieds d'arachide, et étaient plus exposées à la prédation. Cela laisserait supposer qu'en milieu réel, l'arachide pourrait constituer une barrière qui, tout en limitant la migration de ces larves, renforcerait leur mortalité dans les bicultures. Ces observations appuient les conclusions de Litsinger et al. (1991). L'hypothèse "des ennemis naturels" prévoit un renforcement de la faune utile dans les polycultures à cause de la diversité de sources nutritionnelles (proies, pollens et nectars) et d'abris qu'elles offrent (Root, 1973; Risch, 1981; Litsinger et al., 1990). On pourrait ainsi supposer que les ennemis naturels présents dans nos parcelles de bicultures y ont renforcé la diminution du nombre de foreurs. Selon les conclusions de l'IRA (1987), le CMS 8602 donne un rendement supérieur à celui des variétés locales. Nos résultats ne montrent d'écart significatif entre celui-ci et le Bafia que sur les monocultures à Yaoundé (Tableau 2). On pourrait donc penser que de conditions agronomiques adéquates (bonne fumure, date et densité de semis appropriées, etc.) pourraient facilement donner à ce dernier une performance comparable à celle du premier. La monoculture de maïs n'a présenté aucun avantage sur la biculture maïs-arachide du point de vue du rendement. Au contraire, ce dernier était significativement meilleur sur certaines bicultures (Tableau 2). Ceci confirme les résultats des travaux de Schilling (1965) et Mutsaers (1976) qui ont relevé des gains de rendement du maïs, du sorgho ou du mil associés à l'arachide, ou au niébé. Selon Giller et al. (1991), Mohapatra et Pradhan (1992) et Reynolds et al. (1993), le transfert horizontal de l'azote fixé par la légumineuse vers la céréale expliquerait cette augmentation du rendement. Nos résultats qui démontrent que la biculture maïs-arachide diminue le nombre de foreurs, s'accordent avec ceux de Waterworth (1994), Coderre et al. (1989), Kfir (1990) et appuient l'hypothèse de la concentration de ressources. RÉFÉRENCES
The following images related to this document are available:Photo images[cs00039b.jpg] [cs00039a.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}