 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
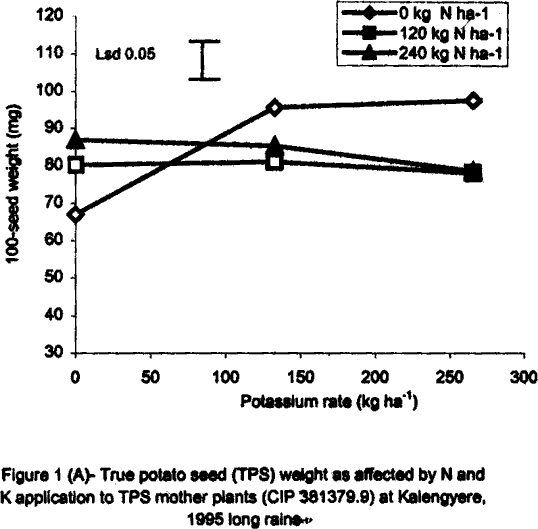
African Crop Science Journal, Vol. 8. No. 4, pp. 375-385 EFFECT OF MOTHER PLANT-APPLIED NITROGEN AND POTASSIUM ON SEED WEIGHT AND seedling EMERGENCE OF TRUE POTATO SEED C.R. KANZIKWERA, D.S.O. OSIRU1, J.S. TENYWA2, E. ADIPALA1 and A.S. BHAGSARI3 Code Number: CS00040 INTRODUCTION One of the major constraints to sowing true potato seed (TPS) as an alternative to planting seed tubers for potato (Solanum tuberosum L.) production is the low seedling vigour during early emergence under field conditions. The genetic constitution of the seed and the environmental conditions it is subjected to before and after harvest, affect its seedling emergence and field establishment (Harrington, 1971; Ross, 1980). Seed production and quality ultimately depend on flower formation, flowering intensity, nutrient flow from the mother plants, preceding fertilisation and formation of the gametophyte. Other environmental factors that may affect seed formation and development include; temperature, light, water and the kind and quantity of available nutrients (Pallais et al.,1987). The production of vigorous seed is generally associated with ideal growth conditions of the mother plants. Conditions that disfavour complete seed development, therefore, result in lack of uniform germination, low seedling vigour and decreased seedling performance under field conditions (Dickson, 1980). Increased assimilate transport towards the developing reproductive structures is required for production of high quality seed (Krauss, 1978). In tobacco, N application to the mother plants, increases seed germination and enhances germination uniformity (Thomas and Raper, 1979). In lettuce, a linear relationship was found between seedling vigour, nitrogen supply and general soil fertility (Soffer and Smith, 1974). Higher yields were obtained from common bean (Phaseolus vulgaris) seed of low weight, but high N content than from heavy seed with low N content (Ries et al., 1970). These findings indicate that, edaphic conditions present during mother plant and seed development can significantly influence the quality of the seed produced. The specific mother plant nutritional requirements for production of high quality TPS have not been systematically investigated. This study was conducted to assess the carry-over effects of N and K application to potato mother plants on TPS quality, in terms of seed weight, seedling emergence and vigour of the seedlings. We have reported elsewhere (Kanzikwera et al., 2001a, b) on the effect of these nutrients on flowering and berry set, and dry matter and nutrient partioning of true potato seed mother plants. MATERIALS AND METHODS Potato mother plants of three potato genotypes, CIP 381379.9 (Kisoro), CIP 800212 and CIP 381403.1, were raised at Kalengyere Research Station (2450 meters above sea level) and Namulonge agricultural and Animal Production Research Institute (NAARI) (1150 masl) during 1995 long rains (LR), 1996 short rains (SR) and 1999 short and long rains. Three levels of potassium: 0, 132.8 and 265.6 kg K ha-1 and nitrogen: 0, 120 and 240 kg N ha-1 were applied factorially in a randomised complete block design, with three replicates at each site. Muriate of potash (KCl) and urea were the K and N sources, respectively. All the K rates and half the rates of N rates were applied into furrows and mixed with the soil before placement of seed tubers. The remaining N halves were top-dressed just before bloom (30 days after planting, DAP). Immediately after application of the second N dose, the urea granules were ploughed into the soil to minimise volatilisation losses. Plot size was 7.0 m x 2.0 m and consisted of three rows, each of fifteen tubers spaced at 1 m x 0.5 m. Uniform size tubers (about 60 g) at about the same physiological age were used as mother tubers. Rutuku, a popular local variety, was planted in separate plots and was used as the pollen source for hybrid TPS produced from a cross between Kisoro (female) and Rutuku (male). The crop was hand-weeded twice and light-hilled at the second weeding. At 45 DAP, pollen was collected from Rutuku plants into vials using a hand-vibrator. Two to three flower buds at late bud or early unfolding stage were selected from Kisoro mother plants in the two middle rows of each plot. They were emasculated by carefully removing the anthers using a scalpel, and were then hand-pollinated in early mornings (8.00-10.00 a.m.). Berries were harvested 5-6 weeks after pollination (WAP), when they just started to ripen. Berries from each plot were collected in polyethylene bags, counted and weighed using a top-loading Harvard Trip Balance (OHAUS). They were then ripened in bags at 27±2 °C for six days. Seeds were extracted onto a 0.5 mm sieve, under running tap-water. They were cleaned and dried at room temperature (27 ± 2°C). The seeds were stored in paper envelopes in an incubator, at about 40 °C for 6 months, to break dormancy (Pallais, 1990). Random samples of 100 seeds from each fertiliser treatment were picked and weighed on a mettler H-80 weighing scale. The hybrid seeds of CIP 381379.9 (Kisoro) x Rutuku were used for greenhouse studies in 1996 and 1999 to assess seedling vigour and emergence, as well as seedling dry matter accumulation. This was done at NAARI. Open-pollinated seeds were not used for the greenhouse studies to avoid compounding genetic and fertiliser treatment effects. Sixty randomly selected seeds (in 1996) and 120 seeds (in 1999) from each fertiliser treatment were sown in wooden trays in two rows of ten seeds each in 1996 and of 20 seeds in 1999, and each of these replicated three times. Seed lots from three treatments were planted in each tray, previously filled with sterilised soil (mixture of sand and peat moss in a ratio of 1:1), at a spacing of 5 cm x 2 cm and a sowing depth of 0.5 cm. In addition, three TPS progenies from the International Potato Centre (Nairobi) namely, IP 88001, IP 88006, and KP 90178.8, were sowed for comparative purposes during 1996. The seed had been raised from mother plants supplied with 500 kg diammonium phosphate ha-1.The trays were watered regularly and uniformly, using tap-water to ensure good moisture supply. Dithane M-45 (Mancozeb 80% WP) was used to control seedling damping-off. Counting of seedling emergence in each treatment was done daily for 21 days. Seedlings were cut at soil surface 30 days after sowing, weighed, oven-dried at 70 °C for 48 hr and re-weighed. The coefficient of velocity of emergence (CVE) was used as a measure of emergence rate. Equation 1 illustrates the procedure for CVE computation. A high CVE value generally indicates that more seeds emerged over a short emergence time (Scott et al., 1984): CVE = 100 [S∑Ni/S∑NiTi] ..... Equation (1) where N, is the number of seeds germinated on day i, and T is the number of days from sowing. Emergence percentage (EP) 10, 15 and 21 DAS was calculated as: EP = 100 [En/T] ................... Equation (2) Where, E = the number of seeds emerged on day n, and T is the total number of seeds planted. Seedling vigour was evaluated at 15 and 21 DAS using a 1 to 5 scale, where: 1 = small seedlings with stunted growth; 2 = moderate growth but visually stunted; 3 = good growth; 4 = vigorous, tall plants with green foliage; and 5 = vigorous, green, strong stem, ready to transplant. The seedling emergence data were arcsine-transformed (Gomez and Gomez, 1984) and analysed using the General Linear Model (SAS, 1987). RESULTS Effect of N and K application on true potato seed (TPS) weight. In the long rainy season of 1995, at Kalengyere, K or N had no significant effect on 100-seed weight (Table 1). However, their interaction significantly (P< 0.01) influenced this parameter (Fig. 1A). In the hybrid seed obtained from CIP 381379.9 (Kisoro) x Rutuku, the effect of K application on 100-seed weight was inversely related to that of N. In other words, increased application of K at low levels of N decreased this parameter and vice-versa.
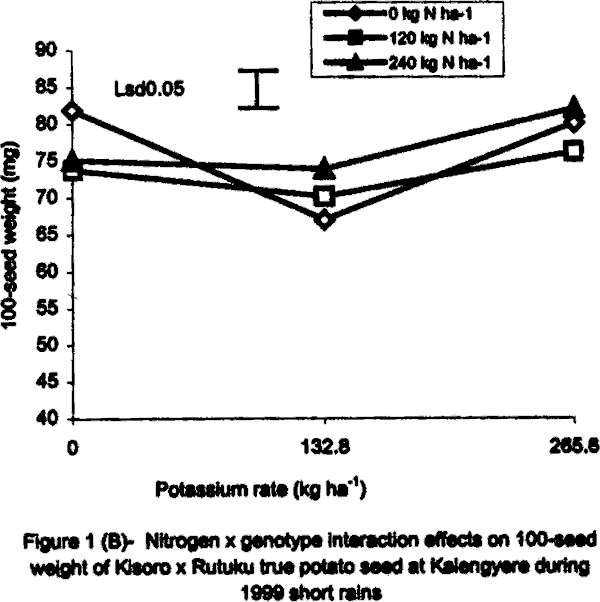
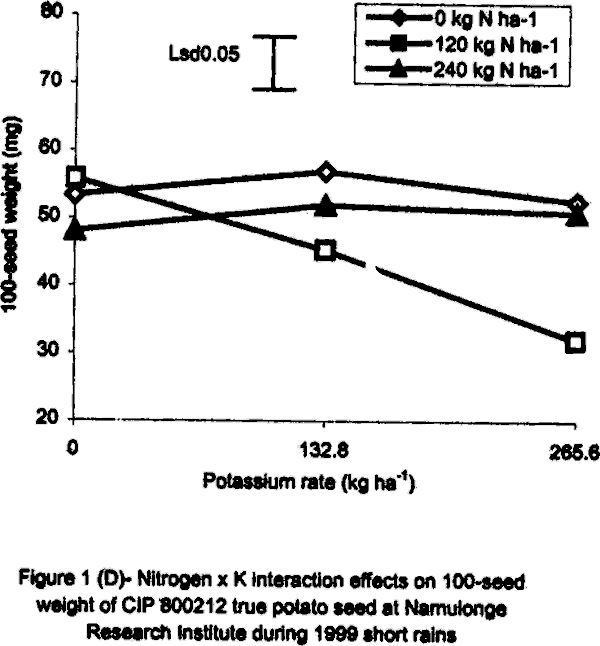
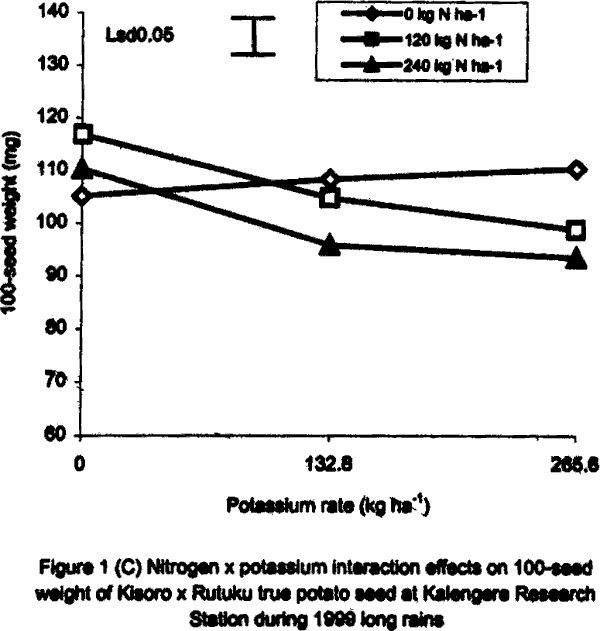
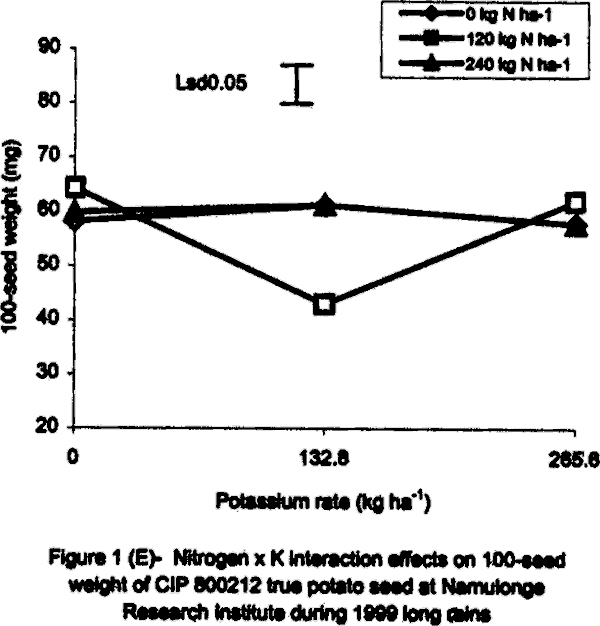
With regard to 1996 SR, there were significant (P<0.05) varietal differences in 100-seed weight. Seeds obtained from open-pollinated (OP) genotype CIP 800212 and hybrid seeds from ‘Kisoro x Rutuku’ crosses were significantly (P<0.05) heavier than OP seeds from CIP 381403.1 (Table 1). In addition, N and K significantly (P<0.05) increased TPS weight. Similarly at Namulonge, seed weight varied significantly (P< 0.05) between potato genotypes. Seed obtained from CIP 381403.1 was 19.4% heavier than that of CIP 800212. Nitrogen and K application, however, had no significant influence on seed weight during the season. It was not possible to produce hybrid seed (Kisoro x Rutuku) at Namulonge because Kisoro variety did not set berries at this location throughout the study period. Its seed weight data are, therefore, not presented. In the case of 1999 SR at Kalengyere Research Station, an inverse relationship was detected between 100-seed weight and number of seeds per berry in CIP 800212, the latter had more but lighter seeds than Kisoro (Fig. 1B). Furthermore, K applied at a rate of 132.8 kg ha-1 produced the highest number of seeds per berry, but with the lowest seed weight (Table 1). However, K rate of 265.6 kg ha-1 significantly (P<0.05) increased seed weight by 21% over the control. Nitrogen application had no significant effect on seed weight neither did the N x K interaction (Fig. 1B). In the case of Namulonge, neither variety nor K had a significant influence (P<0.05) on seed weight. However, there was a significant N x K interaction on 100-seed weight (Fig. 1D). Potassium rates higher than 132.8 kg ha-1 depressed seed weight, while lower rates increased the parameter. There was a negative interaction between N and K on this parameter in that, their joint application led to a decline in seed weight (Fig. 1D). Joint application of K at rate of 132.8 kg ha-1 and N at rate of 120 kg ha-1 decreased seed weight the most. During 1999 LR, TPS weight was quite high compared to earlier seasons of the study, and ranged from 96.8 to 113.3 mg in CIP 800212 and Kisoro, respectively (Table 1). However, neither variety nor fertiliser main treatment significantly influenced this parameter. Nitrogen x K interactions were significant (P<0.01) on seed weight. Potassium alone increased seed weight, while joint N and K application depressed it indicating a negative interaction between the two nutrients (Fig. 1C). Seed weight was neither influenced by N nor K by the main effects at Namulonge. However, there were significant (P<0.01) N x K interactions on this parameter (Fig. 1D). Similar to 1999 SR, joint application of N at 120 kg ha-1 and at 132.8 kg K ha-1 depressed seed weight the most. Nitrogen application at 240 kg ha-1, without K application was not significantly different from the control (0 N kg ha-1) (Fig. 1E). Seedling emergence. In the 1996 greenhouse experiment, seedling vigour varied significantly (P<0.05) among the TPS progenies, ranging from visually stunted seedlings to vigorous ones (Table 2). The vigour was generally higher in the local hybrid seed than in the imported one. Application of K and/or N significantly (P<0.05) reduced seedling vigour. Application of the highest K rate (265.6 kg ha-1) produced the most stunted and weakest seedlings, which had consistently lower emergence than the other rates (Table 2). On the contrary, N application improved seedling vigour. Seedlings from imported seed produced under a heavy fertilisation regime (500 kg DAP ha-1) had intermediate vigour but were visually stunted. Seedling dry weight did not differ significantly among treatments in both study years (1996 and 1999) (Tables 2 and 3). However, joint application of N and K significantly (P<0.05) depressed seedling dry matter accumulation (Table 3).
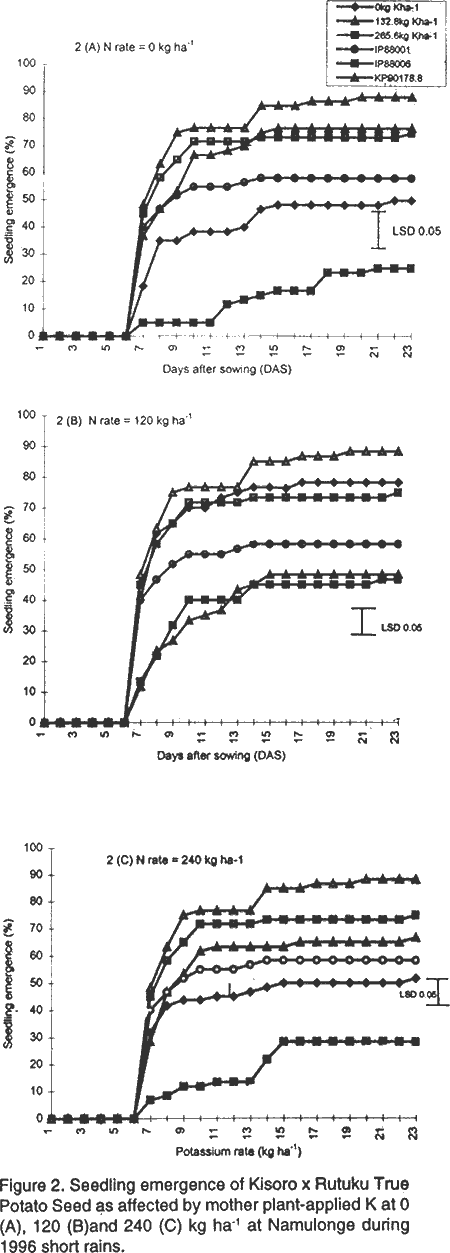
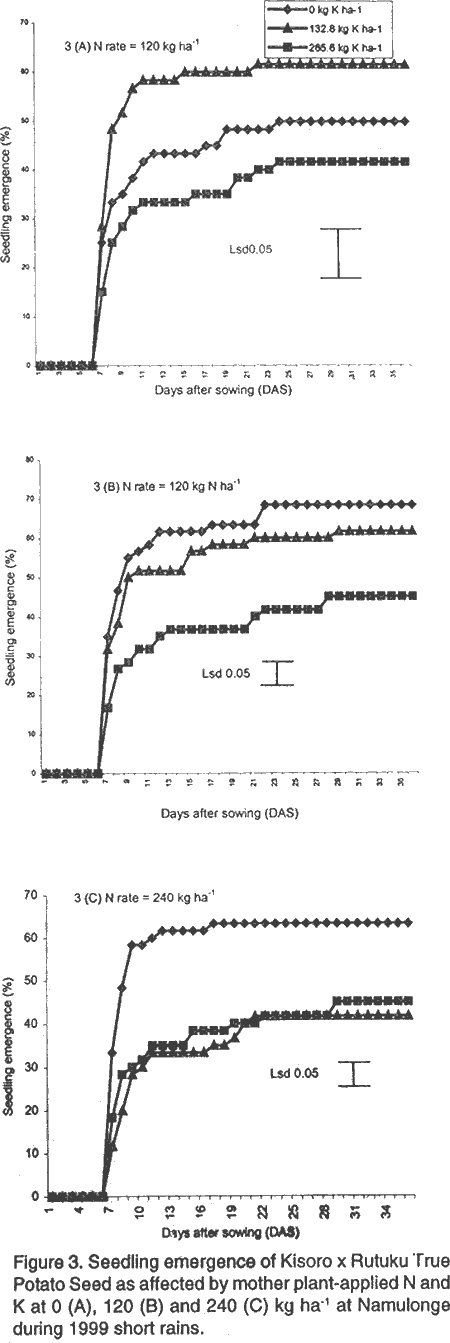
Vigour and dry matter accumulation in potato seedlings as influenced by mother plant-applied N and K at Namulonge. During 1996, there were no significant differences in seedling emergence between the local and imported hybrid TPS progenies as measured by CVE (Table 2). Potassium and N application also had no significant influence on this parameter. The imported progenies, KP 90178.8 and IP 88006, were outstanding in seedling emergence at 10, 15 and 21 days after sowing. The performance of seeds raised under fertilisation regime consisting of 132.8 kg K and 120 kg N ha-1 was quite comparable to and not significantly different from that of the imported TPS progenies. Similarly, during 1999, both N and K significantly (P<0.05) increased seedling emergence (Table 3). Imported progenies were not available for evaluation during the year. Seedling emergence trends are shown in Figure 2 and Figure 3. Without N application (Fig. 2A), the lowest rate of emergence was achieved at the 265.6 kg K ha-1 regime and was highest at 132.8 kg K ha-1 where the emergence was quite comparable to that of the imported seed. When K and N were applied jointly, the rate of emergence declined significantly and reflected a negative nutrients interaction. Sole application of nitrogen at 120 kg ha-1 produced seed with a high rate of seedling emergence, which compared closely with that of the best performing imported progeny (Fig. 2B). At the N rate of 240 kg ha-1, application of K at 132.8 kg ha-1 resulted in seed with significantly (P<0.05) higher seedling emergence than the 265.6 kg ha-1 rate (Fig. 2C). Locally produced hybrid seed, whose mother plants were supplied with 120 kg N ha-1 or 132.8 kg K ha-1, had similar emergence results to those of IP 88006 and significantly (P<0.05) higher emergence than IP 88001 produced from mother plants supplied with 500 kg diammonium phosphate ha-1. Seedling emergence trends in 1999 were very similar to those of 1996. Potassium application at 132.8 kg ha-1 had the highest seedling emergence in the absence of N (Fig. 3 A), while joint N and K application significantly (P<0.05) suppressed the parameter (Fig. 3B and 3C). DISCUSSION From this study, K application rates higher than 132.8 kg ha-1 were associated with low 100-TPS weight. It was also observed that the highest rate of K (265.6 kg ha-1) produced seedlings with the lowest emergence and seedling vigour. Similar relationships between seed size and/or seed weight and germinability and vigour were observed by Delouche (1980) and Pallais et al. (1987). In fact, seed weight has been proposed as a character for selecting high yielding TPS progenies (Dayal et al., 1984). The poor field performance of TPS produced from mother plants supplied with high levels of K, therefore, could be attributed to the low 100-TPS weight produced at these K levels. Mother plants supplied with 120 kg N ha-1 or 132.8 kg K ha-1 consistently produced seed that had the highest field performance among the locally produced TPS. However, a combination of K and N produced seed of poor quality. Potassium and N application evidently had negative interaction on seedling emergence. This negative interaction was also observed on seed weight of hybrid TPS in this study, and elsewhere on maize (Karlen et al., 1987). These findings are in agreement with the suggestion that high TPS weight increases seedling vigour and sowing value (Dayal et al., 1984; Pallais et al., 1987). The beneficial effects of N to the mother plants has been related to its role in delaying senescence and, therefore, allowing berries enough time to receive assimilates and mature on the mother plant (Delouche, 1980; Marschner, 1995). Fox and Albrecht (cited by Delouche, 1980) associated rapid germination and vigorous seedlings with high protein content in wheat seed. High protein content, in turn, was associated with N fertilisation. Lopez and Grabe (1973) reported that application of high N levels to wheat plants increased protein content of the seeds from 6.6 to 10.8%. They further observed that seeds with high protein content germinated faster and developed into larger seedlings compared to average seeds with higher dry matter content. However, increase in seed protein content was accompanied by reduction in seed size and reduced starch content of the endosperm. They concluded that the increase in seed protein due to high N may be restricted to the protein storage species in the endosperm and does not affect embryo and membrane proteins. Abdul-Baki (1980) noted that these endosperm protein reserves are important during seedling growth, especially in N deficient soils. Since the increase in endosperm protein content is associated with low endosperm starch content (Lopez and Grabe, 1973), any increase in protein content raises the density of the seed by altering the protein/starch ratio. Hence, the trait of high seed density, with high protein and low starch content, may be the same factor that determines high seed vigour. In studies conducted in Lima, the amount and rate of germination of TPS of different weights were found to be similar although plant size at transplanting was significantly larger for heavy seed than light seed (Malagamba, 1988). In conclusion, whereas the mechanisms by which N may affect seedling vigour have been reported, knowledge is still lacking as to how K affects seed germinability, seedling emergence and vigour. The results of this study, however, clearly indicate that K increased TPS sowing value and that either N or K could be used to improve field performance of TPS at the study site. In the present study, N and K rates of 120 and 132.8 kg ha-1, respectively, were best for production of TPS with high seedling emergence and vigour. ACKNOWLEDGEMENT This work was done with the support of National Agricultural Research Organisation (NARO) and United States Agency for International Development (USAID)-Washington for a collaborative research on Solanum potato between Makerere University and Fort Valley State University, Georgia, USA. REFERENCES
The following images related to this document are available:Line drawing images[cs00040c.gif] [cs00040g.gif] [cs00040a.gif] [cs00040f.gif] [cs00040b.gif] [cs00040e.gif] [cs00040d.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}