 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 8. No. 4, pp. 387-402 EFFECT OF SOURCE, TIME AND METHOD OF NITROGEN APPLICATION ON GROWTH AND YIELD COMPONENTS OF POTATO IN KENYA G.K. GATHUNGU, S.I. SHIBAIRO, S.M. GITHIRI, M.W.K. MBURU, P.S. OJIAMBO1 and H.M. Kidanemariam1 (Received 20 July, 1999; accepted 24 March, 2000) Code Number: CS00041 INTRODUCTION Low soil nitrogen (N) is known to be a factor limiting potato production in Kenya and many parts of the world (Harris, 1992; Reckle et al., 1997), especially when there is adequate water supply (Goh and Haynes, 1986). Nitrogen fertiliser increases the leaf area which increases the amount of solar radiation intercepted and consequently, increases dry matter production by different plant parts (Krishnappa, 1989). It increases potato yields by increasing the number of tubers formed (Kotsyuk, 1995), duration of tuber bulking, and nitrogen uptake by the plant (Martin, 1995). Nitrogen is a key element in growth and development of crop plants and it influences the yields mainly through leaf area expansion, crop development, crop quality, and susceptibility to lodging (Addiscott et al., 1992). Nitrogen is one of the most abundant elements in plants, constituting 0.5-5 percent of the plant dry weight (Lindblad and Guerrero, 1995) and is a constituent of all proteins, many metabolic intermediates, and of nucleic acids (Goh and Haynes, 1986; Salisbury and Ross, 1991). Potatoes are known to be heavy feeders of nitrogen (Singh, 1995; Veeranna et al., 1997). Low N application rates, improper timing of fertiliser application and faulty application methods (De Datta et al., 1983) are some of the factors which lead to low potato yields. When moisture is not limiting, N is applied before planting or at planting and between 2-4 weeks after emergence or as a split application (before or at planting and 2-4 weeks after emergence). The application methods of the fertiliser vary from broadcast, in bands or placing fertiliser near the seed tuber. In East Africa, Diammonium Phosphate (DAP), Calcium Ammonium Nitrate (CAN), and sulphate of ammonium (SA) are important N sources, and the fertiliser can be applied either as placement in the planting furrow or broadcasting (Ngugi, 1982). Other sources of N such as Urea, Ammonium Sulphate Nitrate (ASN), ammonium nitrate and CAN are also used depending on their local availability. The recovery of N by the crop is usually low (Legg and Meisinger, 1982) and much of the non-recovered N is lost through denitrification, immobilisation, or by leaching out of the root zone (Keeney, 1982). The various N fertilisers used contain either the nitrate N (NO3-) or the ammonium N (NH4+). The NO3- is subject to leaching (Cameron and Haynes, 1986) and NH4+ to volatilisation (Haynes and Sherlock, 1986). Hence, these fertilisers could have different effects on growth and yield of potatoes owing to different availability to the potato plant. There is thus a need to establish ways through which such losses can be lowered and, consequently, increase the efficiency of agronomic use of these fertilisers. However, relationship between the time and method of application of the various sources of N fertilisers on growth and yield of potatoes under Kenyan conditions has not been documented. The objective of this study was to determine the effect of time and method of application of different sources of N fertiliser on (a) growth and development, (b) yield and yield components, and (c) N content of potato leaves and tubers. MATERIALS AND METHODS Seed of potato cultivar Dutch Robjin used in this study was kindly supplied by the National Potato Research Station based at Tigoni. The potatoes were grown at Kabete field station, University of Nairobi between December 1997 and March 1998 (experiment I) and between April and August 1998 (experiment II). The site has well drained, deep, dark reddish brown to dark red friable clay with humic topsoil (Humic Nitisols) derived from Kabete trachyte (Nyandat and Michieka, 1970). The soil pH ranges between 5.2 and 7.2 in topsoil and 5.2 and 7.7 in subsoil. Available K, Ca, Mg, and P ranges from low to fairly high levels. Total soil N is about 0.26 %. A granular form of triple super phosphate (TSP) containing 46% P2O4 was broadcasted over the furrows to provide the recommended rate of 230 kg P205 ha-1 (AIC, 1981) and then covered with a thin layer of soil to minimise direct contact between the fertiliser and seed tubers. The treatments consisted of three ammonium sulphate nitrate, ASN sources of N, Urea (46% N), CAN (26% N), and ASN (27% N), each to provide 150 kg N ha-1 (Sikka, 1994). For each of the source, three times of N application were used; single early application at planting (EA), split application with half at planting and the rest at 5 weeks after emergence (SA), and single late application 5 weeks after emergence (LA). For each of the N sources, two application methods were used; broadcast and placement at 0.05 m from seed crop within the furrow. The experiments were laid out in a randomised complete block design in a split plot arrangement, and replicated three times. Sources of N constituted the main plots and a factorial combination of methods of application and the times of application constituted the sub plots. Each experimental plot measured 3 m x 3 m. The plant spacing was 0.3 m x 0.75 m within and between rows, respectively. In experiment I potato were hand seeded on 16 December 1997 and on 31 March 1998 in experiment II. Seed tubers were placed in furrows above the fertiliser, which had been covered with little soil to prevent scorching and then covered by a thin layer of the soil. The seeds were partially covered to enable location of the seed row in the furrow and subsequent application of N treatment after planting.The crop received 776.2 mm and 586 mm of rainfall for experiments I and experiment II, respectively, and no extreme temperatures were recorded (Table 1). Supplemental irrigation was done immediately after planting in both experiments. The recommended normal agronomic practices for potato production were followed in each experimental season.
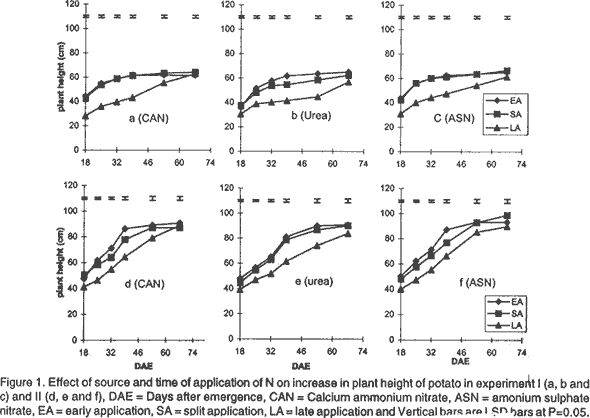
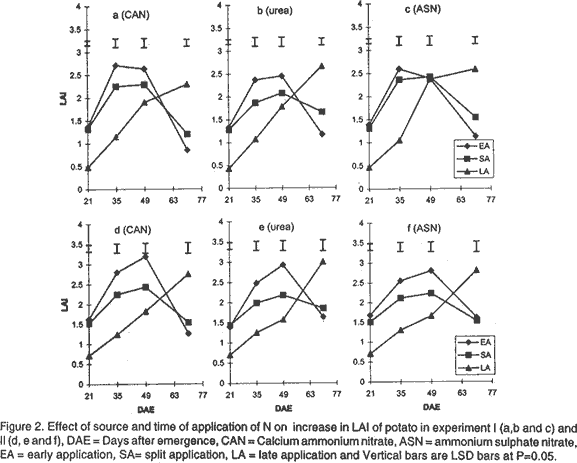
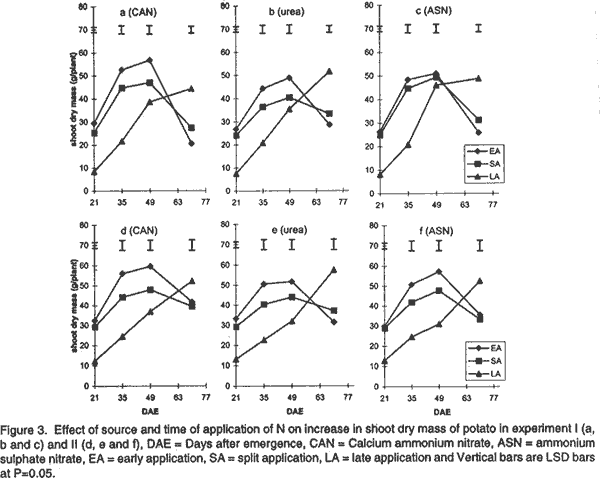
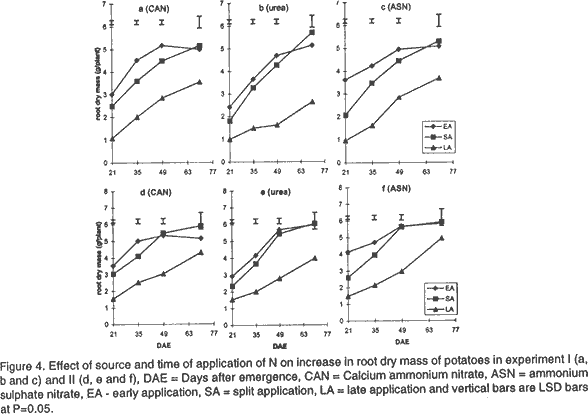
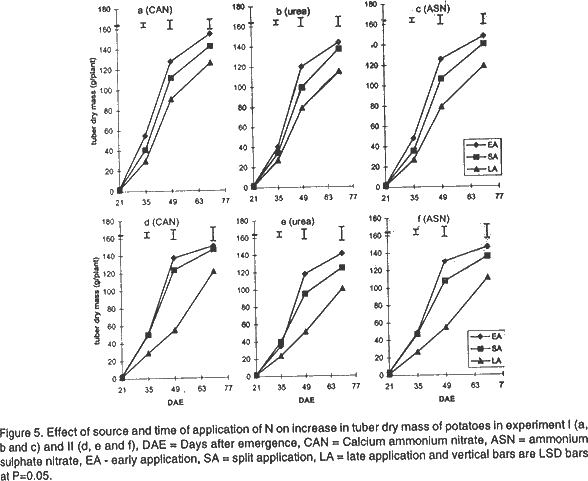
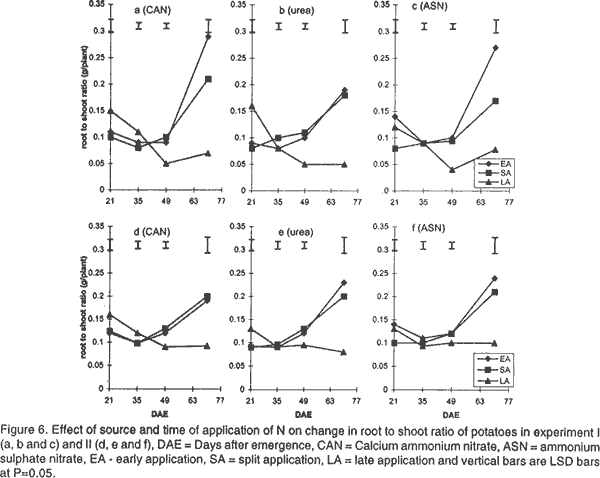
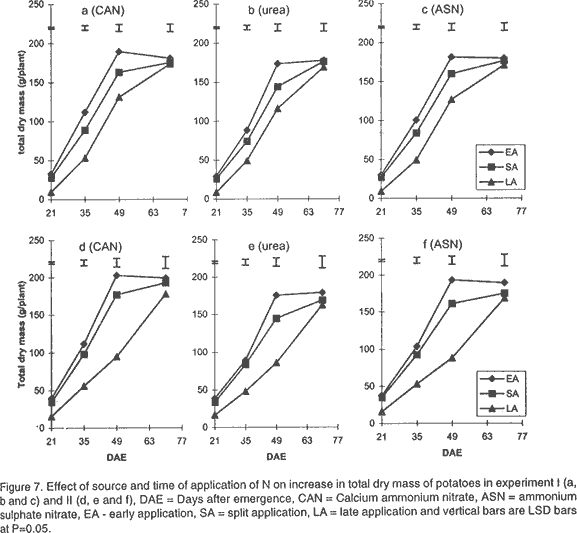
Effect of N on potato growth and development. Plant heights were measured at 18, 25, 32, 39, 53 and 67 days after emergence (DAE) in both experiments. Plant height was determined on three plants randomly selected from the middle three rows. A meter rule was used to determine the height of the plants when all branches were held upright. The Leaf Area Index (LAI) of potato leaves was determined on 15 fully expanded leaves selected from a sample of 3 potato plants randomly selected from each plot. Using a cork borer, 1-cm diameter discs were excised from the leaves, put into 0.164 M x 0.164 M envelopes. The remaining leaf portions and the envelopes were then dried in an oven at 80°C. After 72 h the dry mass of the leaf discs (Lw discs) and the total dry mass of the leaves (LW) were measured. The LAI was calculated using the relationship: LAI = [LW x (LA discs /Lw discs)] x n, where: LA discs = Leaf area (m2) of the discs, Dry mass of the shoot and root systems were determined at 21, 35, 49, and 70 DAE in both experiments using three plants, which were randomly selected from the middle rows in each plot. Potato shoot system comprised of all plant parts above ground level, while the root system comprised of the roots, stolons, and tubers dug from the ground. The shoot and root systems were separately placed in sampling papers and transported to the laboratory. Dry mass of the leaves, stems, roots and stolons was determined by drying in the oven (Model number TV80UL 508032, Memmert, Germany) at 80°C for 72 h. Tubers were cut into smaller pieces, placed in the sampling papers and dried at 105°C for 72 h to obtain tuber dry mass. Total dry mass was obtained by adding shoot dry mass to the root dry mass. Effect of N on potato yield and yield components. The potatoes were hand harvested from the middle three rows at 112 and 119 days after planting in experiment I and II, respectively. Harvesting was done by carefully digging out the potatoes with a hoe from the soil followed by the removal of the haulm. The tubers were then put in separate paper bags. Fresh tubers of ten plants from each plot were used to determine the number of tubers per plant and tuber yield per plant. Determination of N levels in leaves and tubers was made on 9 potato plants for each treatment at 80 DAE and 98 DAE, respectively. The leaves were oven dried at 80°C for 72 h. To facilitate faster and uniform drying the tubers were cut into smaller pieces before being oven dried at 105°C for 72 h. The dried leaves and cut pieces of tubers were separately ground using a dry mill and the powder placed in paper bags which were left in the oven at 72°C. Nitrogen was then determined using 20 mg of each sample by Kjeldahl method (Hind, 1995). Statistical analysis. Repeated measures analysis was done on height change in all the experiments using SYSTAT statistical software (Wilkinson et al., 1992). Shoot and root dry mass, number of tubers and yield/plant and tissue N content (data transformed and percent N calculated as in Annon. (1979) before analysis) were analysed using general linear model (GLM) procedures with S as the main plot, and T and M as split plots. Treatment means were separated using Fisher’s Least Significance Difference (LSD) procedure at P = 0.05. RESULTS Effect of N on potato growth and development. Plant height differed significantly among the N sources and time of application of N at the various growth stages of the potato. Source of N had no significant influence on plant height in experiment I (Fig. 1). However, taller plants were observed in potato that received N as ASN during early and split application in experiment II. Plant height was greatest where N was applied early or as a split application and lowest in the late applied N up to 53 DAE. There was no significant difference in the plant height among the potato 67 DAE in both experiments. LAI significantly differed among the sources and time of N application during the different stages of growth. Potatoes that received N as CAN or ASN had significantly higher LAI than those that received urea when N was applied early or as a split application between 21 and 35 DAE in experiment I and upto 49 DAE in experiment II (Fig. 2). In contrast, potatoes that received N as urea had low LAI than those receiving CAN or ASN for both early and split applied N between 21 and 49 DAE and the highest LAI at 70 DAE. LAI persistently increased between 21 and 35 DAE and decreased at 70 DAE in early and split application of N in both experiments. LAI persistently increased with growth in late applied N potato. LAI was more in the late applied N potatoes, than in the early or split applied N at 70 DAE in both experiments. LAI of the late split applied N was significantly greater than that of early applied N at 70 DAE in experiment I. However, there was no significant difference between split and early applied N at 70 DAE in experiment II. Shoot dry mass increased between 21 and 49 DAE and decreased at 70 DAE in both experiments (Fig. 3). Highest shoot dry mass was observed in potatoes that received N as CAN followed by those that received ASN in early or split N application between 21 and 35 DAE in Experiment I and up to 49 DAE in Experiment II unlike those that received urea. However, potatoes that received urea had high shoot masses at 70 DAE, for both early and split applied N in experiment I. Shoot dry mass persistently increased upto 70 DAE in late applied N potatoes. Shoot dry mass was more in late than in early or split applied N at 70 DAE. Low shoot dry mass 70 DAE in the early or split applied N was as a result of the withering of the stems and leaves. Split applied N potatoes had high shoot dry masses than the early applied N potatoes 70 DAE in experiment I. However, there was no difference in shoot dry masses between split and early applied N plants 70 DAE in experiment II. Root dry mass increased with growth in all potatoes in both experiments (Fig. 4). Significant differences in the response of root dry mass to different sources of N were not observed. Potatoes that had early and split applied N had higher root dry masses than those that had late applied N. Root dry mass in the crop with late applied N was significantly lower than that of early or split applied N at all growth stages. Compared to split applied N, potatoes with early applied N had higher root dry masses between 21 and 35 DAE. However, there was no difference in root dry mass in potatoes that received early and split applied N at 49 and 70 DAE in both experiments. Tuber dry mass increased in all potatoes with growth in both experiments (Fig. 5). Potatoes that received early application followed by split application of N as CAN or ASN had the highest tuber dry masses at all stages of growth. Significantly lower tuber dry masses were observed with urea. Potatoes treated with split applied N had less tuber dry mass than early applied N. Potatoes grown in plots where N was applied late had the lowest dry masses. Root to shoot ratio differed with source and time of application of N at each stage of growth in both experiments (Fig. 6). The root to shoot ratio was highest in potato which received N early, followed by those with split application of N 70 DAE in both experiments for all sources of N. There was no significant difference in root to shoot ratio in potato treated with different sources and times of application of N at 35 and 49 DAE. When N was applied as CAN and urea the potatoes that received N late in the growth season had the highest root to shoot ratio 21 DAE. Potato that had late application of N had the lowest root to shoot ratio 70 DAE. Total dry mass differed significantly with source of N and time of application of N with growth in both experiments (Fig. 7). Potatoes which received CAN had the highest total dry masses at all times of application, while those that had urea had the lowest total dry masses at all growth stages. Total dry mass was highest with early followed by split application of N and lowest with the late application of N for all sources of N. The total dry mass increased between 21 and 49 DAE in potatoes where N was applied early in growth unlike in the split and late applied N where it increased up to 70 DAE. At 70 DAE, there was no significant difference in total dry mass among the different times of application in experiment I. The method of application and the interaction between the source, time and method of application of N had no significant effect on stem height, shoot and root dry mass, LAI, root to shoot ratio, tuber and total dry mass (P < 0.05). Effect of N on potato yield and yield components. The ratio of tuber to the total dry mass content (TDM) differed significantly (P<0.05) among the sources and times of N application at 70 DAE (Table 2). Highest ratio of tuber to total dry matter was observed in potatoes that received N as CAN followed by those with ASN and lowest with potatoes which received N as Urea. Early N application, followed by split applied N had the highest ratio of tuber to total dry matter. Conversely, low ratios were observed with late applied N.
Number of tubers per plant was not significantly affected by the source of N in both experiments (Table 3). However, the time of application of N significantly influenced the number of tubers per plant. The number of tubers were higher in plots treated with early, followed by split N application. Late applied N treatment had the lowest number of tubers per plant. Plants in experiment II had less number of tubers per plant than those in experiment I.
The potato tuber yield significantly (P<0.05) differed amongst the sources and times of application of N in both experiments (Table 3). Application of CAN followed by ASN resulted in high tuber yield. Urea application, however, resulted in low tuber yield. Early followed by split N application led to the highest tuber yields. However, late N application had the lowest potato tuber yields. Overall plants in experiment II had higher tuber yields (kg plant-1) than those in experiment I. The method of application and the interaction between the source, time, and method of N application had no significant effect on tuber to total dry mass 70 DAE, number of tubers and tuber yield per plant. Leaf and tuber N content was not affected by the source of N but it significantly (P<0.05) differed with different application times of N in both experiments (Table 4). Late application of N led to high leaf N than in the early and split application of N. Tuber N was only affected by source of N in experiment I but not in experiment II. In experiment I, potatoes which received CAN had the highest, while those with Urea and ASN had the lowest tuber N. There was no significant difference in tuber N content between tubers that received N either as Urea or ASN. Late and split N application of N led to high tuber N in both experiments. Potato that received early N had the lowest tuber N. The method of application and the interaction between the source, time, and method of N application had no significant effect on leaf and tuber N content.
DISCUSSION Effect of N on potato growth and development. Increase in the dry mass of plants depends on the amount of photoassimilates fixed through photosynthesis (Lawlor, 1990). Solar radiation interception, water supply, CO2 availability, air/soil temperature, and mineral nutrients are factors that determine the amount of photosynthesis (Salisbury and Ross, 1991; Kormondy, 1996). Since K is not limiting in Kenyan soils (Ngugi, 1982), lack of P and N have been responsible for low crop yields observed in potatoes. Although, CAN, ASN, Urea, DAP among other fertilisers are used in potatoes for the provision of N, the extent to which they affect growth and development is not well studied. In this study, plants treated with CAN followed by ASN showed larger LAI, higher root and shoot dry masses, total dry masses and taller plants than those treated with urea. The fertilisers CAN and ASN proved superior to Urea. These fertilisers had N in the form of NH4+ and NO3- ions compared to urea mainly with the NH4+ N. The superiority could be attributed to the fact that, a mixture of NO3- plus some NH4+ ions produces more growth than either of the ions provided alone. Under field conditions NH4+ is known to stimulate tuber swelling (Osaki et al., 1995) while the NO3- stimulates branching of stolons, and promotes shoot growth of potatoes. CAN proved superior to ASN probably due to the presence of Ca2+ ions in it. Ca is important in the maintenance of cell membrane integrity and in cell division, and hence affects plant growth and development greatly. Urea resembles NO3--N in its susceptibility to leaching and NH4+-N in transformations following hydrolysis (Broadbent et al., 1958) and, therefore, it is subject to losses through volatilisation. Such losses may explain the observed low dry matter accumulation, and hence, lowered plant growth and development in potatoes treated with Urea in this study. The timing of application of fertiliser N is an important factor in determining the rate of growth and yield of a plant. The key to plant growth and development depends on the establishment of a large LAI that is durable through the reproductive phase, achieved through adequate N and water supply. In this study, greater LAI, shoot dry mass, root dry mass and plant height were observed in potatoes (during the early growth stages upto 49 DAE) that received early followed by the split application of N especially when the source of N was either CAN or ASN. The results further show a low root to shoot ratio in potato with the early and split application of N earlier in the growth season, indicating that most of the photoassimilates were channelled towards the production of the shoot rather than the roots. Potatoes receiving early and split applied N showed more shoot production by having taller potatoes with high LAI, leaf dry masses and shoot dry masses. Hence, early application of N followed by split application of N enabled a high interception of solar radiation, mainly due to the greater photosynthetic surface area of the crop (LAI). This resulted in increased photosynthetic capacity and supply of the assimilates leading to increased growth. The greater growth in early or split application of N may have resulted into faster root growth, increase in bulking rate, and high root dry mass. A high total root dry mass suggests that early and split fertiliser application facilitated early root growth. Early root growth enables faster shoot growth rates since there is greater capture of other nutrients resulting from increased root surface area of absorption. Apart from affecting root growth, increased growth may have led to more total dry matter accumulation and translocation into tubers, and hence, leading to high tuberisation and tuber dry mass yields in the potatoes treated with early and split applied N (Gunasema and Harris, 1968). It has been suggested that split application of fertiliser leads to increased growth and hence improved yield over applying the fertiliser whole at planting (Kidin and Zamaraev, 1996). In this study potatoes that received split application of N showed lower shoot dry mass, LAI, leaf dry mass, and total dry mass between 21 and 49 DAE than those that received early application of N. This suggests that the growth that occurs is mostly manifested in maintenance of vegetative phase, early in the growth season with very little concomitant increase in tuber growth. Due to the additional supply of N at the second split application, there is increase in growth but this too is more manifested in vegetative than root growth. These results therefore show that potato growth and development will be improved most by single application at planting than through split application of N. Late application of N, especially as Urea, resulted in low LAI, shoot and root dry masses, and low root to shoot ratio during early growth of potatoes. The low LAI led to low interception of solar radiation and hence low photosynthetic capacity to support growth in the early growth stages. Consequently, these potatoes showed inferior heights, root and tuber dry mass development. A low root to shoot ratio late in growth of potatoes that received late N showed that more shoot growth was supported than root growth. Furthermore, the results show that no difference in total dry mass was observed 70 DAE. This was contributed mainly by the shoot dry mass since the root dry masses and the tuber dry masses were lower in potatoes that received late application of N. This suggests that application of N late in the season led to increase in vegetative growth at the expense of tuber growth, and hence low potato growth and development occurred. The availability of applied N to the potato crop depends on the method of fertiliser application (Addiscott et al., 1992). The method of application of N (broadcast or placement in furrow) or its interaction with source or time of application did not affect growth and development in this study. It has been suggested that in a situation where there is no moisture stress, the method of application of N could be of less importance except under conditions of low soil fertility (Neubauer, 1993). A total of 776.2 mm and 586 mm of rain were received for experiment I and II, respectively. This translated to 8.26 mm/day and 3.83 mm/day for experiment I and II potatoes, respectively. Rainfall amounts greater than 3 to 5 mm/day is considered high for potato growth (Manrique, 1984). The rains in this study therefore led to high soil moisture and hence differences between the methods of application of N could not be observed. However, it is possible that under moisture stress conditions, the mobility of N may be interfered with and hence differences in methods of application of N may arise. There is therefore, need to undertake studies to determine the effect of moisture stress and method of application of N on growth and development of potatoes. The present results, however, suggest that application of CAN or ASN as a single application, early in the growth period will improve growth and development. Urea application does not lead to similar improvements in growth and development. Split and late application of N only leads to delayed growth and development and hence may not be adapted for increased yields in potatoes. Effect of N on potato yield and yield components. Increase in potato yield is mainly due to increase in number and size of the potato tubers (Kotsyuk, 1995). The number and size of tubers depend on the rate of tuber initiation and the amount of photoassimilates generated through photosynthesis. N has been shown to affect tuber initiation and the rate and amount of photosynthates (Kormondy, 1996; Salisbury and Ross, 1991). This study showed that both source and time of application of N affected potato tuber yield and the leaf and tuber N. CAN had the highest tuber yield followed by ASN and urea, respectively. The number of tubers per plant was not affected by source of N. The results therefore, suggests that the source of N as either CAN, ASN or urea had no effect on tuber initiation and hence the number of tubers per plant of these potatoes. This suggests that the observed differences in yield were explained by differences in tuber mass per plant. Although tuber mass per plant was not recorded in this experiment, it is likely to have been highest with potatoes that received CAN followed by ASN compared with Urea. This is supported by results of the observed high tuber dry mass of potatoes supplied with CAN followed with ASN and low with urea at 70 DAE. High tuber dry masses which led to high yields per plant in potato that received N as CAN are likely to be due to the accumulation of photoassimilates, that were high in these potato due to an earlier establishment of a greater LAI as discussed above. In addition, it is more of the result of translocation of photoasssimilates towards the tubers than the shoots in these potatoes. The advantage of using CAN or ASN over Urea was attributed to the provision of N both as NH4+ and NO3- ions as discussed above. The disadvantage of using Urea was attributed to losses through leaching and volatilisation (Broadbent et al., 1958). The low growth and development observed in potatoes supplied with Urea resulted to plants with lower masses and hence lower yields. The results of this study show that time of application of N affected potato yield significantly. High potato yields were obtained in potatoes that had early followed by split and late application of N. As discussed above, early application of N led to high rates of growth and development resulting in high LAI and generally high shoot and root growth. The increase in root growth resulted in increase in both the number (bulking rate) and size of tubers. A high root to shoot ratio 70 DAE in potatoes which received early application of N, suggests that most assimilates were channeled towards tuber growth than vegetative growth. Martin (1995) suggested that N supply increases the duration of tuber bulking and this may result in large sized potato tubers hence high yields. In this study, early application of N may have led to early tuber initiation. The tubers therefore, had a long bulking period resulting in large sized tubers and hence more yield. Potatoes that received split application had lower yields than those receiving early application of N. It is possible that the lower growth rate observed early, followed by the later fast growth rate late in the season did not support increase in tuber number or sizes of the potatoes. This therefore, suggests that the added N only supported vegetative growth rather than tuber growth and size. It has been argued that establishment of a high LAI and leaf area duration early in the growth will increase the photosynthetic capacity and the amount of photosynthates produced and, consequently, greater production (Kormondy, 1996). Conversely, a late establishment of the vegetative growth will lead to low photosynthesis hence low production. In this study, the potatoes which received late application of N showed lower potato yields. This occurred since the plants had the fewest tubers per plant. In addition these potatoes had the lowest tuber dry mass and tuber to shoot ratios. It is therefore suggested that the potatoes receiving late N supports more vegetative growth than tuber growth. These potatoes therefore showed lower yields. In experiment I potatoes had more tubers per plant but less yield than those of experiment II. Conversely, experiment II potatoes had less number of tubers per plant but more yield per plant than those of experiment I. Though the data on tuber size was not recorded in this study, the lower yields in experiment I suggests that the tubers were small sized than those in experiment II. Adequate soil moisture supply in experiment I could have favoured tuber initiation resulting in more tubers. Cool temperatures, in the later growth stages, favoured more tuber bulking resulting into large sized tubers and consequently high yields per plant in experiment II. The method of application of N did not affect tuber number and yield per plant. It is therefore concluded that either of the two methods of application of N can be adopted with CAN applied as a single application at planting. Also, application of CAN or ASN as a single application, early in the growth period will improve yield and yield components of potatoes. Urea application does not lead to similar yield improvements in potato. Split and late application of N only leads to delayed growth and development and hence may not be adapted for increasing yields in potatoes. In this study the source of N did not significantly influence the concentration and the quantity of N in the leaves or tubers. This probably indicates that though the rate of absorption of the form of N may be different, the distribution of the N in the plant is not influenced by the source of N. Low N content in leaves and tubers was observed in potatoes that received early application of N. However, late application of N resulted in the highest leaf and tuber N content. High supply of N has been reported to increase the N content in leaves and tubers (El -Gamal, 1996; Veeranna et al., 1997). It is possible that where N was applied early in the growth season, the plants root system was not fully developed hence much of the applied N was not absorbed and stored in the plant system. More N was, therefore, lost through leaching from the soil and through senesced leaves falling from the plant. In late application of N, the crop had a well developed root system and therefore most of the applied N was absorbed into the plant system. In addition, in late application of N most of the growth had ceased and therefore, the N supplied was not used in the growth processes resulting in being stored in tissues. The leaves, however, had a greater percentage of N content than the tubers. It has been suggested that the leaves are the first sinks for the N supplied before it is translocated to the tubers (El-Gamal, 1996), and that may explain the results of this study. The decline in N content is interrupted or delayed by additional dose of N in the season (Vitosh and Silva, 1996). In this study additional dose of N in the split-application of N increased the N content in both the tubers and leaves over the treatments where the fertiliser was applied early in the growth season. The method of application of N did not significantly influence leaf and tuber N content in this study. As indicated earlier this could be as a result of similar access to N by the potato because the nutrient was equally distributed within the furrow when either of the two methods was used. At the time of application of N soil moisture was not limiting and therefore N was equally mobile in the furrow whether it was broadcasted or placed. However, it is possible that under moisture stress conditions, the mobility of N may be interfered with and therefore differences may arise. There is, therefore, need to undertake studies to determine the effect of moisture stress and method of application of N on leaf and tuber N content of potatoes. It is recommended for better growth and development and, consequently, yields of potatoes. ACKNOWLEDGEMENTS The authors thank International Potato Centre (CIP), Sub-Sahara Africa region, Nairobi, Kenya for providing the research funds. We are also grateful to Mr. Charles Lung'aho (NPRC-Tigoni) for providing seed material used in this study. REFERENCES
The following images related to this document are available:Line drawing images[cs00041e.gif] [cs00041a.gif] [cs00041d.gif] [cs00041c.gif] [cs00041b.gif] [cs00041g.gif] [cs00041f.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}