 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 9. No. 2, pp. 339-350 EMBRYOGENESE SOMATIQUE A PARTIR D’EMBRYONS IMMATURES DE BLE TENDRE: EFFET GENOTYPIQUE K. MZOURI, M. AMSSA et E. H. BOUIAMRINE (Received 4 March, 2000; accepted 2 November, 2000) Code Number: CS01015 INTRODUCTION Dans un programme de sélection in vitro, le choix des génotypes qui pourraient être ultérieurement améliorés dépendra principalement de leur aptitude à la culture in vitro. Celle-ci réside dans leur capacité à produire des cals embryogènes et de régénérer par la suite des plantes. Chez le blé tendre, espèce d’une grande importance économique à l’échelle mondiale, plusieurs travaux de régénération à partir de divers explants (embryons, inflorescences, feuilles...) ont été rapporté (Maddock et al., 1983; Redway et al., 1990; Barcelo et al., 1991; Rajyalakshmi et al., 1991; Menke-Milczarek et Zimny, 1992a, b; Ponitka et al., 1993; Barakat, 1994). Cependant, les meilleurs résultats pour l’induction des cals embryogènes et la régénération ultérieure, sont obtenus à partir d’embryons immatures (Ozias-Akins et Vasil, 1982; He et al., 1986; Rotem-Abarbanell et Breiman, 1989; Bouiamrine, 1998). Par ailleurs, la capacité de régénération des plantes à partir d’embryons immatures est fortement influencée par la composition du milieu de culture (Mathias et Simpson, 1986; Carman et al., 1987b; Fekete et Pauk, 1989; Purnhauser, 1991; Borrelli et al., 1992; Zair et al., 1995), le stade de développement des explants (He et al., 1988; Qureshi et al., 1989) et surtout du génotype (Van Sint Jan et al., 1990; Chowdhury et al., 1991; Felföldi et Purnhauser, 1992; Menke-Milczarek et Zimny, 1992b; Hagio, 1994; Dusautoir et al., 1995; Bouiamrine, 1998). La présente étude a comme but d’évaluer la capacité d’embryons immatures chez 20 variétés de blé tendre à former des cals embryogènes et à régénérer des plantes. Par la suite, ont été déterminées les différentes corrélations existantes entre les paramètres étudiés (taux de callogenèse, taux de germination zygotique, taux de cals embryogènes, taux de régénération et le nombre moyen de plantules par cal régénérant). MATERIEL ET METHODES Vingt variétés de blé tendre (Triticum aestivum L.) ont été testées pour leur aptitude à la culture in vitro: Nasma, Tegyey 32, Potam, Marchouch, Jouda, Sibara, Saïs, Acsad 59, Saba, Kanz, Achtar, Baraka, Khair, Saada, Tillila, Massira, Rajae, Amal, Mehdia et 1765. Ces variétés, inscrites au catalogue officiel marocain à l'exception d’une variété: 1765, étaient choisies pour leurs caractères agronomiques adéquats. La semence de ces variétés était fournie par l'I.N.R.A. (Institut National de la Recherche Agronomique). Les explants utilisés dans le cadre du présent travail consistaient exclusivement en embryons immatures. Les semis étaient réalisés en lignes en plein champ. Les épis étaient prélevés 2 à 3 semaines après l'anthèse. A ce stade, les embryons ont une taille de 1 à 1,5 mm. Pour chaque variété, des caryopses étaient débarrassés de leurs glumelles et désinfectés sous agitation continue pendant 20 à 30 secondes dans de l’éthanol 70°C; puis 15 minutes dans l’eau de Javel 30%. Les caryopses étaient, par la suite, rincées trois fois dans l’eau distillée stérile (sous flux laminaire). Après cette étape de désinfection, les embryons étaient prélevés stérilement, sous loupe binoculaire, et placés, la face scutellaire vers le haut, dans des boites de Pétri (10 cm de diamètre) contenant le milieu callogène. Le milieu utilisé était MS additionné de 2 mg l-1 de 2,4-D (Tableau 1).
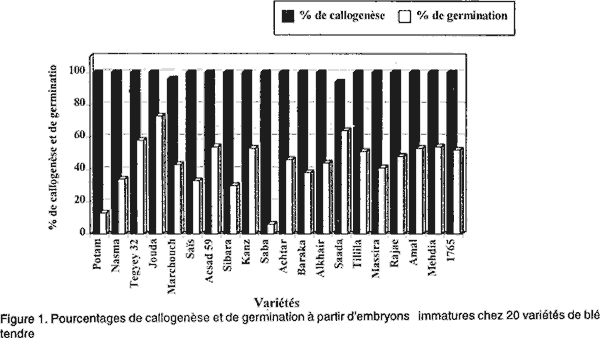
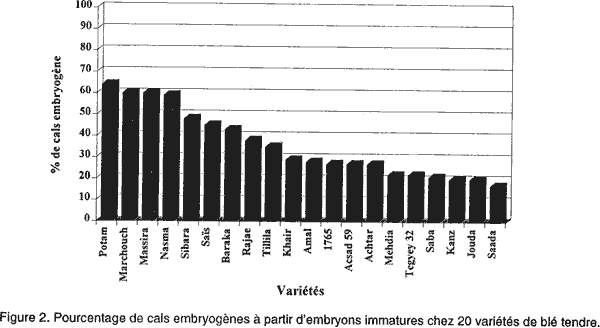
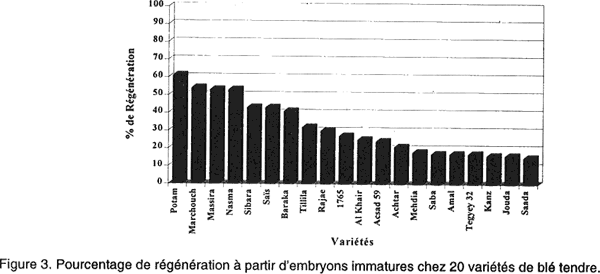
L'ensemencement se faisait à raison de 12 embryons/boite de Pétri. Ces boites étaient scellées au parafilm. Pour induire les cals, les cultures étaient incubées à l'obscurité dans une salle climatisée maintenue à 25°C ± 2°C. Les cals en régénération étaient soumis à une photopériode de 16 heures de lumière/24 heures. Les plantes régénérées, après 1 mois (Redway et al., 1990), étaient repiquées sur un milieu de croissance R2 (Tableau 1). A partir de ce moment, les plantes étaient repiquées régulièrement toutes les quatre semaines, sur ce même milieu frais afin de les maintenir vivantes jusqu’à ce qu’elles atteignent une hauteur de 5 cm et un système racinaire bien développé. Ensuite, ces plantes étaient transférées au sol. Pour l'évaluation de l'aptitude à la callogenèse et à la régénération à partir des cals issus des embryons immatures de 20 variétés de blé tendre, nous avons considéré les paramètres suivants: • l’induction des cals, mesurée par le taux d’explants ayant produit du cal, après 4 semaines de culture in vitro et le pourcentage de germination zygotique, déterminé par le nombre d’embryons mis en culture ayant germé sur le milieu de callogenèse; • la capacité des embryons mis en culture à former des cals embryogènes, mesurée par le pourcentage de cals embryogènes [(nombre de cals embryogènes/nombre total de cals de la variété) x 100]; • le pourcentage de régénération [(nombre de cals ayant régénéré des plantules/le nombre total de cals de la variété) x 100] après 4 semaines de culture sur milieu de callogenèse et deux mois sur milieu de régénération R1; • et le nombre moyen de plantules/cal morphogène estimé après deux mois de culture sur milieu R2. RESULTATS Induction des cals et germination zygotique. Les observations effectuées après quatre semaines de culture ont permis de distinguer des cals compacts, entièrement (photo 1) ou partiellement (photo 2) nodulaires qualifiés d’embryogènes (Nabors et al., 1983; Ryschka et al., 1991) et d’autres non embryogènes qui sont d’aspect translucide et mucilagineux (photo 3). La callogenèse est observée chez la totalité des explants mis en culture (Fig. 1). Cependant, le pourcentage de germination zygotique varie entre 13 % (variété Potam) et 73% (variété Jouda). Taux de cals embryogènes. La Figure 2 illustre les taux de cals embryogènes obtenus à partir d’embryons immatures chez les 20 variétés testées. Un effet génotype très marqué a été observé dans la production des cals embryogènes. Les variétés Potam, Marchouch, Massira, Nasma, Sibara, Saïs et Baraka présentent les pourcentages de cals embryogènes les plus élevés (41% à 64%). Les variétés Rajae, Tillila, Khair, Amal, 1765, Acsad 59 et Achtar présentent des pourcentages intermédiaires (entre 27% et 38%). Alors que les variétés Mehdia, Tegyey 32, Saba, Kanz, Jouda et Saada sont considérés les moins embryogènes (< 22 %). Aptitude des cals à régénérer des plantes. Sur le milieu de régénération, les deux types de cals répondent différemment: les cals embryogènes régénèrent, dans la plupart des cas et donnent des plantules dans 2 à 3 semaines (photo 4). Cette régénération se manifeste dans un premier temps, par une caulogenèse, suivi par la suite d'une rhizogenèse. Mais, dans certains cas, ces cals montrent à leur surface des structures verdâtres, dont la croissance est arrêtée, qui font penser à une inhibition de la germination (photo 5). Les cals non embryogènes montrent parfois des racines avec ou sans chlorophylle (photo 6), mais dans la plupart des cas, aucune morphogenèse n’est observée et les cals tendent plutôt à se nécroser (photo 7). Pour maintenir le développement des plantules régénérées sur le milieu R1, on procède au repiquage sur un milieu sans régulateurs de croissance (R2). Les photos 8 et 9 montrent une régénération à partir des cals placés pendant 4 semaines sur le milieu R2. Les pourcentages de régénération observés (Fig. 3) sont également affectés par le génotype. Les variétés Potam, Marchouch, Massira, Nasma, Sibara, Saïs et Baraka présentent les taux de régénération les plus élevés (41% à 61%). Les faibles taux de régénération (15% à 18%) sont observés chez les variétés Mehdia, Saba, Amal, Tegyey 32, Kanz, Jouda et Saada. Nombre moyen de plantules/cal régénérant (NMPR). Nous avons procédé à l'évaluation du nombre moyen de plantules régénérées par cal régénérant pour les 20 variétés testées. Cette évaluation a eu lieu juste avant leur transfert au sol. Le comptage a porté sur 20 à 25 cals pris au hasard (Tableau 2). Le NMPR est déterminé par le nombre de plantes régénérées d’une variété/le nombre de cals morphogènes de la même variété.
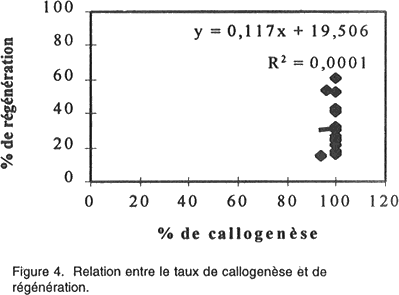
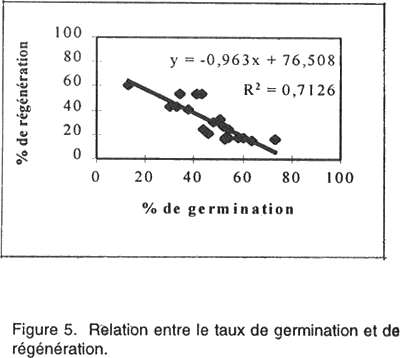
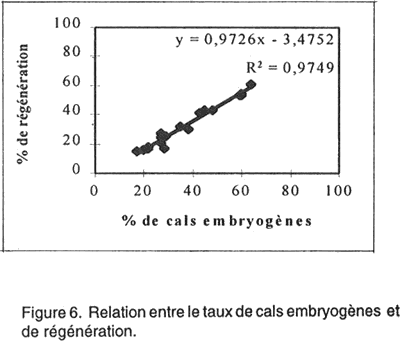
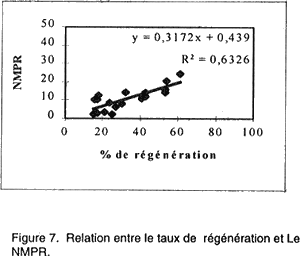
Le rendement en plantules par cal régénérant est également un paramètre affecté par le génotype. Le NMPR varie de 2,1 chez la variété Saada et 24,7 chez la variété Potam. Corrélations entre paramètres étudiés. Pour quantifier l'aptitude des 20 variétés à la culture in vitro, quatre corrélations ont été déterminées. Les résultats présentés par les Figures 4, 5, 6, 7 nous a permis de mettre en évidence: • l’absence d’une corrélation entre le taux de callogenèse et de régénération (R2 = 0,0001); • l’existence d’une corrélation hautement significative (R2 =0,713) entre le pourcentage de germination zygotique et le taux de régénération. Ces deux paramètres sont inversement proportionnelles; • l’existence d’une corrélation très hautement significative (R2 = 0,975) entre les pourcentages de cals embryogènes et de régénération. Ces deux critères sont proportionnelles; • l’existence d’une corrélation hautement significative (R2 = 0,633) entre le taux de régénération et le nombre moyen de plantules/ cal morphogène. DISCUSSION L'embryogenèse somatique désigne l'ensemble des événements provoqués artificiellement in vitro et conduisant à la formation d'un embryon à partir d'une ou plusieurs cellules somatiques. Ce phénomène s’accompagne de prolifération cellulaire qui nécessite la présence de 2,4-D, auxine indispensable dans le milieu de callogenèse. Cette auxine agit comme un signal inductif pour le déclenchement de l'activité prolifératrice de l’explant mis en culture qui donne naissance, par la suite, à un cal. Le 2,4-D contribue également au développement d'embryons somatiques de plusieurs espèces (Vasil, 1987). L’observation des cals obtenus nous permet de distinguer deux types de cals: (1) des cals embryogènes où l’on distingue la formation d’embryons somatiques avec leurs deux pôles: un pôle racinaire et un pôle caulinaire comme chez les embryons zygotiques (ces cals sont caractérisés par un haut pouvoir de régénération); et (2) des cals non embryogènes (compacts lisses ou bien mucilagineux humides) qui, prolifèrent rapidement en donnant naissance, dans certains cas, à des néoformations (tiges, feuilles ou racines) et que l’on qualifie d’organogènes. Ces observations concordent avec celles rapportées par Ho et Vasil (1983) et Botti et Vasil (1983). Ces deux types de cals ont été fréquemment décrits à partir de différentes sortes d’explants aussi bien chez le blé (Nabors et al., 1983, Chowdhuty et al., 1991; Bouiamrine, 1998) que chez d’autres espèces céréalières (Vasil et Vasil, 1981; Bhaskaran et Smith, 1989; Ryschka et al., 1991). Le passage par cal est une étape primordiale en sélection in vitro, parce que ce tissu végétal est une source importante de variabilité génétique (variation somaclonale) et peut être considéré comme un stock du matériel génétique (El Bayoudi, 1991). Par ailleurs, les investigations que nous avons entreprises montrent que la totalité des explants mis en culture ont formé un cal. Des résultats similaires sont obtenus par Maddock et al. (1983), Dogmi (1994) et Bouiamrine (1998). Par contre, l’effet génotype est observé clairement dans l’aptitude à la production de cals embryogènes et à la régénération de plantes chez le blé tendre. Cet effet a été mis en évidence par plusieurs auteurs (Maddock et al., 1983; Carman et al., 1987a; He et al., 1986, 1989; Ou et al., 1989; Felfôldi et Purnhauser, 1992). Ce phénomène ne se limite pas seulement au blé tendre; il a été observé, aussi, chez d’autres céréales, avec des intensités différentes, comme le maïs (Rhodes et al., 1986), l’orge (Hanzel et al., 1985; Lühr et Lörz, 1987), le riz (Fatokum et Yamada, 1984; Van Sint Jan et al., 1990) et le blé dur (Bennici et al., 1988; Dogmi, 1994; et Bouiamrine, 1998). Chez toutes les variétés testées nous avons observé que le taux de régénération est plus ou moins faible par rapport au taux de cals embryogènes. Cette diminution de la régénération s’expliquerait par la présence d’une compétition entre les embryons somatiques du même cal pour l’espace, milieu de culture, etc. Dans ce cas, la fragmentation du cal a permis la régénération de la majorité des embryons somatiques. Une telle diminution pourrait aussi résulter de la variation du nombre de chromosomes (aneuploïdie et polyploïdie), des réarrangements chromo-somiques (délétion, duplication, inversion, translocations, etc.) et des modifications au niveau de l'ADN mitochondriale et chloroplastique (Ahloowalia et Sherington, 1985) causés par des mutations au niveau du génome des cellules méristématiques de l’explant en donnant naissance à des embryons somatiques anormaux qui ne peuvent pas régénérer. C’est ainsi que parmi les 20 variétés de blé tendre que nous avons testées, seulement 7 (Potam, Machouch, Massira, Nasma, Sibara, Saïs et Baraka) ont présenté une aptitude élevée à la culture in vitro. Ces résultats sont en accord avec des travaux réalisés sur d’autres céréales qui ont montré que seulement quelques génotypes sont capables de répondre favorablement aux stimulants morphogénétiques in vitro. Par exemple, chez Hordeum vulgare L., seulement 6/91 génotypes ont formé un cal compact embryogène (Hanzel et al., 1985); alors que, chez Oryza sativa L., 7/31 variétés ont donné des taux de callogenèse, de croissance et de régénération relativement élevé pour être utilisés dans des programmes ultérieurs de sélection in vitro (Van Sant Jan et al., 1990). Chez Sorghum bicolor, seulement 5/250 variétés ont montré une capacité relativement élevée pour la formation des cals et la régénération des plantes à partir d’embryons immatures (Hagio, 1994). L’absence de corrélation entre les taux de callogenèse et de régénération montre que ces paramètres sont contrôlés par des gènes différents. L'indépendance de ces deux caractères a été déjà signalée chez le blé (Sears et Deckard, 1982; Lazar et al., 1983; Chowdhuty et al., 1991), ainsi que chez l'orge (Komatsuda et al., 1989). Les gènes qui contrôlent l’induction d’un cal ne sont pas les mêmes que ceux qui contrôlent sa capacité embryogène et par la suite son aptitude à la régénération. La germination précoce des embryons immatures mis en culture montre une corrélation négative avec la régénération des cals issus de ces embryons. Ces résultats concordent avec ceux réalisés par Felfôldi et Purnhauser (1992) qui ont travaillé sur 44 variétés de blé tendre et 3 variétés de triticale et dans les mêmes conditions que celles appliquées dans cette étude. Ce phénomène peut être expliqué: une telle germination épuise le milieu de culture et retarde le développement normal du cal et par la suite des embryons somatiques. Nous avons observé aussi une corrélation positive hautement significative entre le pourcentage de cals embryogènes et le taux de régénération et une autre entre le pourcentage de régénération et le NMPR. Ces résultats sont en accord avec ceux réalisés sur le blé dur (Bouiamrine, 1998). Les variétés ayant une grande aptitude à produire des cals embryogènes, leur capacité à régénérer des plantes est donc aussi relativement élevé ainsi que leur NMPR. Les pourcentages de germination zygotique, de cals embryogènes, de régénération et le NMPR semblent être des bons indices de l’aptitude à la culture in vitro. L'existence de trois corrélations significatives entre ces quatre pourcentages permet de quantifier l'aptitude des 20 variétés testées à la culture in vitro. Chez le blé tendre, le contrôle génétique de l’embryogenèse somatique et la régénération a fait l’objet de plusieurs travaux (Mathias et Fukui, 1986; Higging et Mathias, 1987; Kaleikau et al., 1989a et b). Des substitutions du stock génétique ont montré que presque tous les chromosomes du génome peuvent intervenir dans la régulation de ces processus (Galiba et al., 1986; Felsemburg et al., 1987; Kaleikau et al, 1989a, b). Néanmoins, un ou deux gènes ont une influence majeure sur la régénération des plantes (Hodges et al., 1985; De Buyser et al., 1992; Henry et al., 1994a et b; Ben Amer et al., 1992a et b; 1995). En plus de l’effet génotype, des facteurs biotiques (conditions de développement de la plante mère, source de l'explant, son stade de développement, sa position sur la plante entière) ou abiotiques (composition du milieu de culture et les facteurs environnementaux) interviennent dans la réponse morphogénétique. L’interaction entre ces facteurs affecte le niveau de la quantité endogène en régulateurs de croissance (AIA, ABA.) des explants mis en culture (Evans, 1984; Rajasekaran et al., 1987; Brown et al., 1989). De tels phénomènes seraient à l’origine des différences dans la réponse obtenue à partir de divers types d'explants de la même plante, ou du même explant cultivé à des stades différents de son développement (Maddock et al., 1983; Vasil, 1987). Une corrélation entre l’âge de l’embryon et la teneur endogène en régulateurs de croissance, particulièrement, l’ABA a été également rapportée (Qureshi et al., 1989). En effet, les embryons à des stades précoces synthétisent suffisamment d’ABA, ce qui explique une stimulation de l’embryogenèse somatique et une diminution du pourcentage de germination. Par contre, les embryons à des stades avancés, contiennent moins d’ABA ce qui favorise leur germination précoce. L’expression de la compétence morpho-génétique in vitro est donc complexe et fortement influencée par les facteurs physiologiques, de développement et epigénétiques, qui influencent la synthèse, le transport et l'utilisation des régulateurs de croissance. Les taux de cals embryogènes et de régénération des cultivars considérés comme récalcitrants (ne répondent pas bien à la culture in vitro) peuvent être significativement améliorés en maximalisant les conditions de cette expression. Cette amélioration de la capacité morpho-génétique chez les cultivars considérés comme ayant une mauvaise aptitude à la culture in vitro, est liée principalement à la mise au point de milieux de cultures adéquats dont les résultats seront présentés dans le prochain manuscrit. References
The following images related to this document are available:Photo images[cs01015h.jpg]Line drawing images[cs01015b.gif] [cs01015d.gif] [cs01015c.gif] [cs01015g.gif] [cs01015f.gif] [cs01015a.gif] [cs01015e.gif] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}