 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 9. No. 2, pp. 385-392 Drought tolerance of some bread wheat genotypes in Ethiopia Desalegn Debelo, Bedada Girma, Zewdie Alemayehu and Solomon Gelalcha (Received 10 October, 1999; accepted 17 December, 2000) Code Number: CS01020 INTRODUCTION The availability of water for biological roles as solvent and transport medium, as electron donor in Hill’s reaction (photosynthesis), and as evaporative coolant is essential for normal plant growth (Bohnert et al., 1995). It may be assumed that all plants have encoded capability for stress perception, signaling and response. But plant species as well as genotypes within a species may vary in sensitivity and response to the decrease in cell water potential caused by drought. The bulk of wheat (Triticum aestivum) production in Ethiopia is under rainfed conditions. A vast wheat production area of the country is characterised by unreliable rainfall both in amount and distribution. Therefore, wheat production in these areas encounters moisture deficit at critical growth stages. This problem has limited the productivity of the crop and its expansion in area in the country. Farmers in some drought prone areas are growing traditional cultivars that are relatively tolerant through adaptation, but are genetically low yielding. Irrigation facilities are not well developed in these areas. Therefore, breeding for drought stress tolerant/resistant varieties remains the principal alternative to increase the productivity of wheat in such environments. Ceccarelli and Grando (1991) argued that progress in yield in stress environments is possible if selection for yield is performed in those environments. According to them, yield under stress can be effectively improved by selecting for yield under this condition, while yield potential can be improved only under non-stress conditions. Since selecting for yield under stress often results in reduced yield potential, it may therefore, be suggested that stable yield under stress and high yield potential may be mutually exclusive. Under moderate stress conditions, however, varieties with high yield potential are still best performers. Ceccarelli and Grando (1991) demonstrated that high yielding varieties of barley were more adapted to favourable growing conditions while the lower yielding landraces were more stable in their performance under stress conditions. Testing and selection of varieties for yield stability under stress sites and years has been considered as an option for improving stress resistance (Ortiz-Ferrara et al., 1991). The nature of drought varies from location to location and year to year with regard to timing and intensity. Consequently, genotype (G) x environment (E) interaction is encountered when varieties are tested across locations or environments. Significant G x E interaction results from the changes in the magnitude of differences between genotypes in different environments. Different stability estimates have been proposed to measure the consistent performance of genotypes tested across a wide range of environments. Pooled analysis of variance combined with joint regression analysis proposed by Yates and Cochran (1938), modified by Finlay and Wilkinson (1963) and made popular by Eberhart and Russell (1966) has been extensively used to study stability. The linear regression techniques, however, were found to confound interaction with main effect (Wright, 1971) and non-linear genotypic response to the environments (Nachit, 1986). To faciliate the partitioning and interpretation of multi-location genotype trials, multivariate methods such as additive main effects and multiplicative interaction (AMMI) procedure with pediction assessment has been developed as a powerful technique (Guach, 1988; Guach and Zobel, 1988). AMMI is a non-conventional analysis of variance which splits the sum of square for interaction into multiplicative components related to eigenvalues (Guach, 1988). The results of AMMI analysis can be presented graphically in the form of biplots in which the cultivar and environment scores are represented by vectors in a space, with starting points at the origin and the end points determined by the scores (Vargas et al., 1999). It is indicated that the distance between end points of any two cultivar vectors is indicative of the amount of interaction between the vectors. Identification of measurable stress resistance traits and determination of their association with yield to use as indirect selection criteria are the critical preludes in breeding for stress tolerance/resistance. Ortiz-Ferrara et al. (1991) observed that increased plant height played an important role under terminal drought stress. This shows that in a breeding programmes intended to develop varieties tolerant to drought stress, careful consideration of plant height enables one to improve drought tolerance. The duration class of genotypes is another important trait in drought tolerance breeding. Nachit and Ketata (1991) showed that grain yields under drought stress were highly related to earliness. To design a wheat variety for a particular adaptation zone, attention must be given to the climate (temperature and moisture) conditions so that the variety will complete its development in a suitable environment (Przul and Mladenov, 1999). Although priority goes to high-yielding varieties, early maturing ones can escape, especially the terminal drought problem. It should be clear that early maturity is not necessarily an exclusive trait associated with drought tolerance of varieties. Thus, the comparative importance of vegetative and reproductive growths have not been emphased in drought tolerance breeding programmes. On the other hand, certain grain characteristics which play important roles, especially in response to terminal drought tolerance/resistance of genotypes, have not yet received adequate attention as a selection criteria. In this study, complementary statistical procedures thought to be useful for drought tolerance breeding were applied to drought stress variety trials conducted in different environments for three years. Linear regression technique and AMMI model were used to evaluate the genotypes tested under drought stress environments for yield stability across such environments. Correlation coefficients were also estimated to evaluate certain pre- and post harvest traits, that may contribute directly or indirectly to yield for their utilisation as an indirect selection criteria for yield under drought stress conditions. MATERIALS AND METHODS This study was conducted in five drought prone locations (Asasa, Dhera, Goro, Kulumsa and Mekelle) in different parts of Ethiopia from 1996 to1998. Mekele and Goro were not included in 1997 and 1998, respectively. Seventeen advanced bread wheat genotypes along with three checks were used at all locations. The trial was laid out in a randomised complete block design with four replicates. Each genotype was drilled on six rows of 2.5 m x 20 cm (3 m2). A seed rate of 150 kg ha-1 (45 g/plot) was used, and planting was done manually. Fertiliser rates recommended for each testing site were applied and nitrogen was split applied (1/3 at planting and 2/3 at tillering). Observations for days to 50% heading and 75% maturity were recorded. Grain filling period was computed by subtracting the number of days to heading from the number of days to maturity. Plant height was measured from ground level to the tip of spikes excluding the awns. The four central rows were harvested manually for grain yield, 1000-kernel weight and test weight determination. The data were subjected to statistical analysis using AGROBASE software. With the intention to identify adaptable varieties to such variable environments, a pooled analysis of variances for grain yield, days to maturity, grain filling period, thousand-kernel weight and test weight was conducted. RESULTS AND DISCUSSION The five testing sites are variable in geographical, edaphic and climatic factors. The three testing years were also different in their seasonal rainfall amount and distribution. (Table 1). The combined analysis of variance for grain yield showed that there were highly significant (P<0.01) differences among the mean squares of the genotypes, the test locations and the G x E interaction (Table 2). The linear component of G x E interaction was also significant (Table 3). This implied that the genotypes responded differently to the variation of the testing environments. Stability analysis using Eberhart and Russell regression model was performed in order to identify adaptable varieties to the different drought prone environments. The result showed that the test varieties HAR 2294 and HAR 2500 had βvalue below unity with very low positive deviation from linearity(0.49%) in their response, indicating their stable yield performance across the stress environments. Varieties such as HAR 2507, HAR 2320 and HAR 2330 showed regression value below unity with moderate to high deviation from linearity of response. Varieties such as HAR 2449, HAR 2501, HAR 2505, HAR 2506 showed beta (β) values above unity with relatively high G x E stability variance (Table 4). This demonstrated that these varieties perform better in relatively favourable environments.
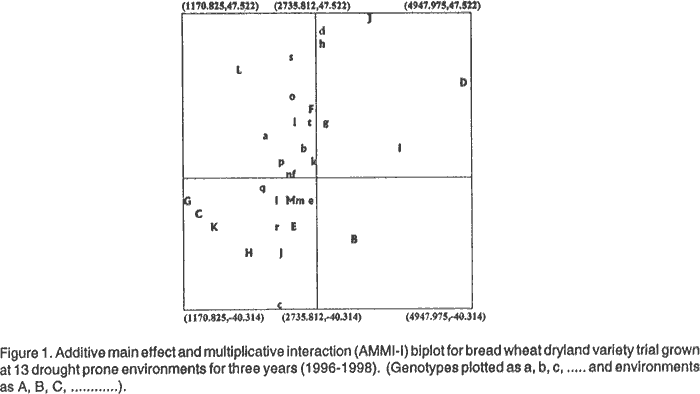
On the other hand, HAR 2508, HAR 2324, HAR 2319 and HAR 2503 showed negative deviation from linearity of response. This indicates that the performance of these varieties does not change much with the change in environment. The superior genotype in grain yield, HAR 2504, had a slope close to unity and low positive deviation from linearity. This showed general stability of the genotype, demonstrating a buffered grain yield performance across stress and non-stress growing environments. Similar result was previously reported for some current commercial varieties (Desalegn et al., 1996). Results of AMMI analysis using the same grain yield data showed that there were highly significant differences among the varieties, the environments and G x E interaction (Table 5). The interaction variance was explained in 13 IPCA. Among these, the first six IPCA axes were significant and explained 93.24% of the interaction variance. This revealed that the attributes accountable for G x E interaction might be roughly grouped into six. These attributes were environmental as well as genotypic by nature.
The three check varieties used had relatively good performance under low moisture stress. Among them, Dereselign and Pavon 76 recorded low positive IPCA axis-1 scores, while K6290-Bulk had low negative IPCA axis-1 score (Table 6). This evidence indicates that the above varieties used as checks have genetic adaptation on the target environments, owing to their low interaction.
The yield potential of the testing sites and the level of discrimination they pose on the genotypes was stratified. The highest mean yield was observed at Kulumsa during 1996 and 1997 (4,947 and 4,306) kh ha-1 whereas, the lowest mean yield was recorded ar Dhera during 1997 (1170 kg ha-1). Asasa and Kulumsa, areas with relatively good rainfall (amount and distribution) showed positive environment IPCA axis 1 scores (Tables 5 and 6). While Goro, Dhera and Mekele, representative of site areas where crop failure is not uncommon, showed negative environment IPCA axis 1 scores. As can be observed from the values of the IPCA axis-1 scores, high seasonal variations were observed at Asasa, Dhera, Goro and Mekele (Table 7). The identification of genotypes that can favourably grow over these variable environments is necessary to attain increased grain yield in these areas.
High variation was observed among the test genotypes not only in grain yield, but also in their interaction across environments (Fig. 1). The highest yield was recorded for HAR 2504, HAR 2505, HAR 2501 and HAR 2508, with a yield range of 3,177 to 3,006 kg ha-1. Among these genotypes, HAR 2504 exhibited low positive IPCA axis 1 score. This indicates that the performance of this genotype changes little with the change in environments. Two out of the high yielding genotypes, HAR 2501 and HAR 2505 had high positive IPCA axis-1 scores, demonstrating their positive interaction with the relatively favourable areas of Kulumsa and Asasa. The other high yielding genotype, HAR 2508, had very low negative genotype IPCA axis-1 score. This implied its little interaction across environments and positive specific interaction effects in the unfavourable environments, Dhera, Goro and Mekele. Most of the test genotypes which yielded higher than the grand mean had positive IPCA axis-1 scores, demonstrating their specific interaction with the favourable environments and their adaptation to such areas (Guach, 1988). Association of some agronomic traits with grain yield. Correlation coefficients were estimated among seven traits including yield (Table 8). All the correlation coefficients were positively or negatively significant except for the association between days to maturity and 1000-kernel weight. All the traits considered showed positive and very strong association with grain yield. This indicates that these traits contributed directly or indirectly to the grain yield under low moisture stress conditions. This suggests that pre-harvest selection of tall plants with longer maturity period enables identification of high yielding varieties. This may be due to stem reserve mobilisation.
Strong positive correlation was observed between grain size and grain yield, demonstrating that the former can be used as an indirect post-harvest selection criterion in yield improvement programmes. Panozzo and Eagles (1999) reported that the rate of grain filling is lower in dry environments than in wet environments. As a result smaller and lighter grains are obtained. However, genotypic variations are obviously detected in the same crop. The association of test weight with grain yield was also strongly positive. Nevertheless, narrow variability was observed for this trait to use it as an indirect selection criterion. In conclusion, both methods of analysis showed that varieties such as HAR 2503, HAR 2508, HAR 2324 and HAR 2504 which showed high mean yield performance and low IPCA axis-1 scores are adaptable across the stress environments. On the other hand, HAR 2501 and HAR 2505 were the best performers in the relatively favourable areas. As yield was positively associated with plant height, late maturity and 1000-kernel weight, these traits can be used as selection criteria for development of tolerant varieties. REFERENCES
The following images related to this document are available:Line drawing images[cs01020a.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}