 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 9, No. 1, March 2001, pp. 109-125
C. R. Kanzikwera, J. S. Tenywa1, D. S. O. Osiru, E. Adipala and
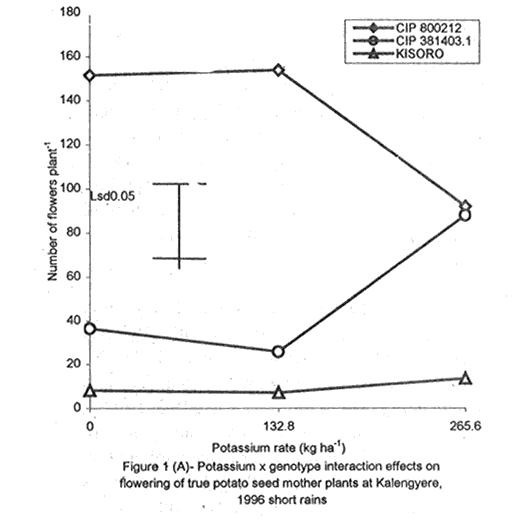
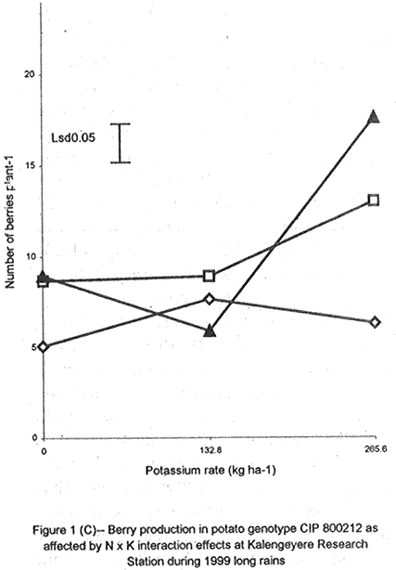
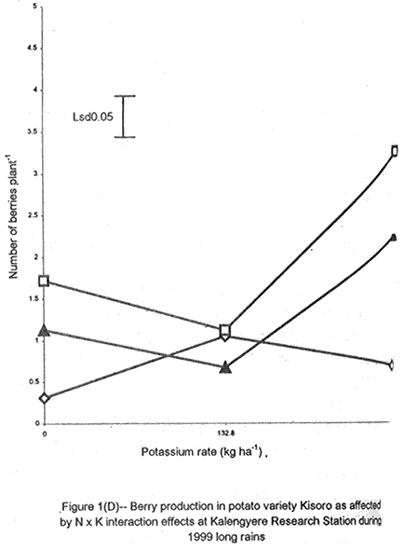
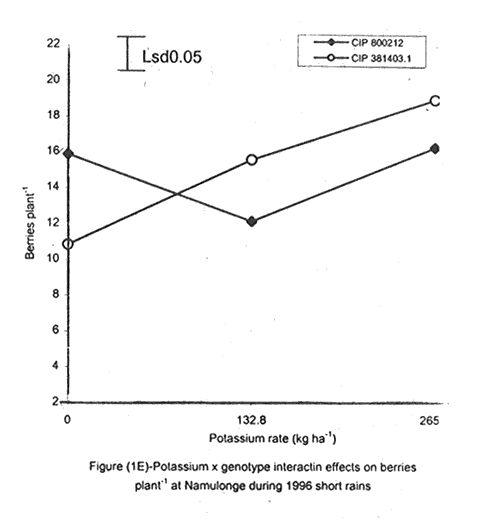
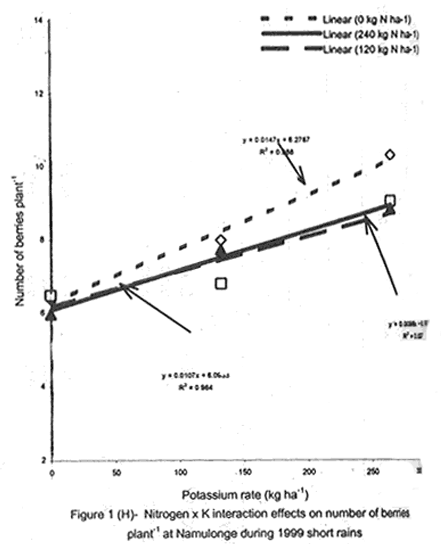
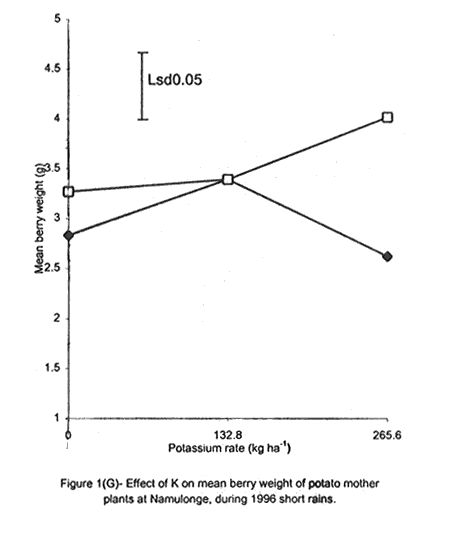
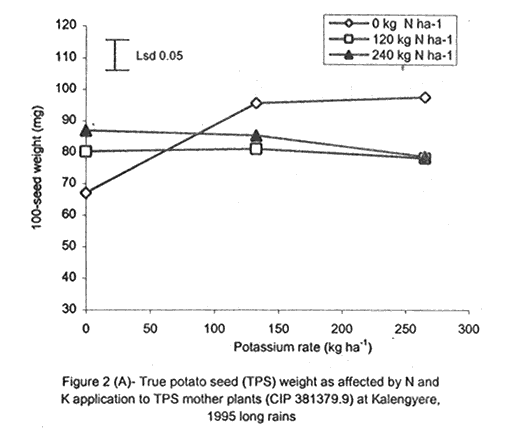
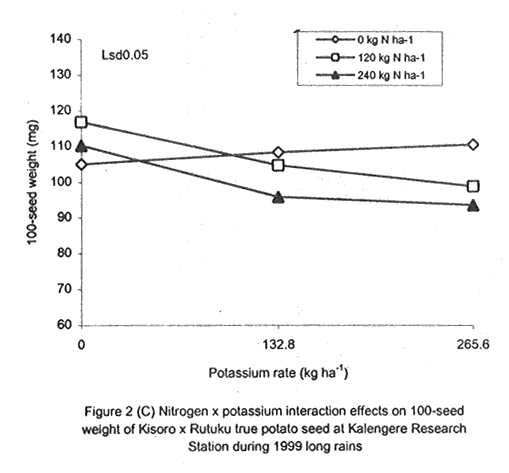
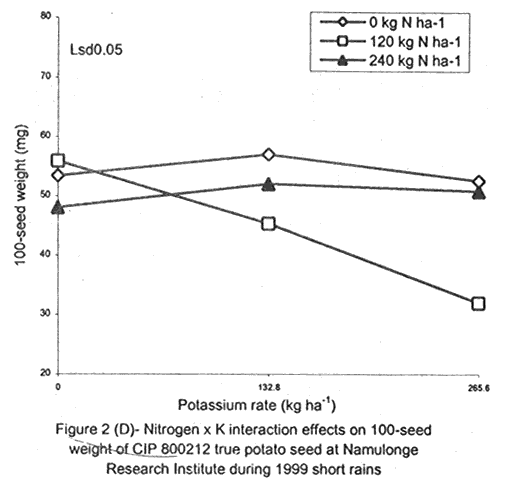
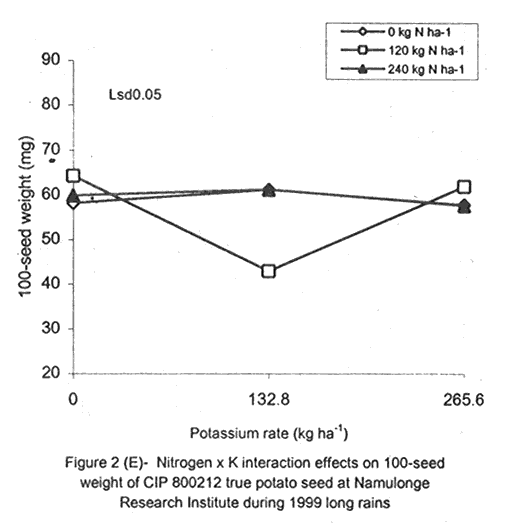
A. S. Bhagsari2 Code Number: CS01041 ABSTRACT Field experiments were conducted at Namulonge Agricultural and Animal Production Research Institute and Kalengyere Research Station during 1995, 1996 and 1999 to assess effect of N and K on flowering, berry set and true potato (Solanum tuberosum) seed yield. Three N (0, 120, 240 kg ha-1) and K (0, 132.8 and 265.6 kg ha-1) rates were applied to true potato seed potato mother plants of three potato genotypes, CIP 800212, CIP 381379.9 (Kisoro) and CIP 381403.1. There was significant (P≤0.05) N x K interaction on flowering. Potassium rate of 266 kg ha-1 increased this parameter more than any other treatment. Mother plants fed with 266 kg K ha-1 produced 26% more berries than the control (0 kg K ha-1). Potassium also significantly (P≤0.05) increased berry weight. Nitrogen application at 240 kg ha-1, reduced berry production. In the hybrid seed obtained from 381379.9 (Kisoro) x Rutuku at Kalengyere, K increased seed weight at low levels of N and depressed it at high N levels. Similarly, N increased true potato seed (TPS) weight at low levels of K and depressed it at high K levels. There was a negative interaction between N and K on this parameter. Number of seeds per berry ranged between 53.7 and 71.2 and was not significantly affected by N application during 1999. Potassium application significantly (P≤0.05) increased number of seeds during the season. Nitrogen and K rates of 120 and 132.8 kg ha-1, respectively, were most optimal in promotion of seed production. An inverse relationship between 100-seed weight and number of seeds per berry was observed. Key Words: Potato genotypes, Solanum tuberosum, true seed, Uganda RÉSUMÉ Des essais en champs ont été conduits à l'Institut de Recherche de production animale et agricole à Namulonge et à la station de recherche de Kalengyere en 19995, 1996 et 1999 pour évaluer l'effet de l'N et du K sur la floraison, l'établissement des baies et sur le rendement des TPS. Trois doses d'azote (0, 120, 240 kg ha-1) et de K (0, 132.8 et 265.6 kg ha-1) ont été appliquées sur les plantes mères des vraies semences de 3 génotypes de pomme de terre; CIP 800212, CIP 381379.9 (Kisoro) et CIP 381403.1. Il y avait une interaction significative (P≤0.05) entre l' N et le K pour la floraison. Le taux de K de 265.6 kg ha-1 a augmenté la floraison plus que les autres traitements. Les plantes mères ayant reçu la dose de 265.6 kg ha-1 ont produit plus de 26% de baies plus que le contrôle (0 kg ha-1) . Le potassium a aussi augmenté significativement le poids moyen des baies. Le taux d'application d'N de 240 kg ha-1 a réduit cependant la production des baies. Chez l'hybride de semence obtenu entre le croisement de 381379.9 (Kisoro) et Rutuku à Kalengyere, le K a augmenté le poids des semences à des niveaux bas d'N et l'a réduit à des niveaux élevés d' N. De même, l'N a augmenté le poids de TPS à des niveaux bas de K et l'a réduit à des niveaux élevés de K. Il y avait une interaction négative entre l'N et le K pour ce paramètre. Le nombre de graines par baie variait entre 53.7 et 71.2 et n'a pas été affecté par l'application de l'N en 1999 LR. L'application du potassium a favorisé significativement (P≤0.05) le nombre de graines durant la saison. Les doses de 120 et 132.8 kg ha-1 d'azote et du potassium respectivement ont été presque optimales dans la promotion de la production des semences. Une relation inverse entre le poids de 100 graines et le nombre de graines par baie a été observée. Mots Clés: Génotypes de pomme de terre, Solanum tuberosum, true seed, Uganda INTRODUCTION The formation and development of seeds strongly depend on both genotypic and environmental factors such as, temperature, water, light and the type and quantity of mineral nutrients available in the soil (Delouche, 1980; Tekrony and Hunter, 1995). Soil N application levels higher than those required for crop production have long been recognised by seed physiologists to improve seed yield and vigour (Delouche, 1980; Gray and Thomas, 1982; Van Staden et al., 1982; Gentry and Below, 1991). Similar levels of N have been reported to increase flower production and pollen germination and is effective in the production of vigorous and high quality TPS (Pallais et al., 1984; Pallais et al., 1987; Pallais, 1991; Maingi et al., 1994). Nitrogen application to potatoes (Solanum tuberosum) before tuber initiation increases the number of tubers per plant and mean fresh tuber weight. Increase in N supply also causes delay in bulking but prolongs the bulking phase and leaf area duration leading to high tuber yield (Sharma, 1990). However, continuous supply of N to plants promotes shoot and root growth, and prevents tuberisation (Ivins and Bremner, 1969; Gunasena and Harris, 1971). These processes are related to the indirect influence of N on the activity and phytohormone balance in the plant, especially, on the levels of gibberellic and abscissic acids and cytokinins (Amzallag et al., 1992). Application of N enhances the export of cytokinins from the roots to the shoots resulting in delayed senescence of the plants. Berries, therefore, have a longer time to mature on the mother plants and a better chance for high quality seed production (Van Staden et al., 1982). Because of the important role N plays in the production of high quality seed, attempts to increase flowering, berry setting and TPS quality without N application may be of little significance (Gupta and Pal, 1989). In our earlier work (Kanzikwera et al., 1997), TPS mother plants showed poor response to K application in flowering, berry set and TPS production. This was attributed to competition between tubers and aerial plant parts for limited assimilates. One way of reducing such competition in potatoes is to ensure that there are adequate assimilates to cater for both vegetative and tuber growth. For TPS production, it may be beneficial to promote other vegetative growth at the expense of the tubers. Application of high rates of N to potatoes, depending on the variety, generally delays tuber initiation and promotes vegetative growth (Marschner, 1990). Poor potato response to K application in flowering and berry set, was also partly attributed to low N and reduced uptake of Mg due to K-Mg antagonism. In order to validate these speculations, a study was conducted to assess interaction effects of K and N on flowering, berry setting and TPS yield. Results on dry matter yield and nutrient partitioning are reported elsewhere in this issue (Kanzikwera et al., 2001). MATERIALS AND METHODS Field experiments were conducted at Kalengyere Research Station (KRS) during the 1995 long rains and at both Namulonge Agricultural and Animal Production Research Institute (NAARI) and KRS during the 1996 short rains and 1999 short and long rains. Soil samples were taken from the sites prior to and after establishment of the study, using standard soil sampling procedures, and analysed for pH, organic matter, soil texture, total N, available P, and exchangeable Na+, K+, Ca2+ and Mg2+ (Okalebo et al., 1993). Site soil characteristics are reported elsewhere (Kanzikwera et al., 2000). Treatments consisted of three levels of K (0, 132.8 and 265.6 kg K ha-1) and N (0, 120 and 240 kg N ha-1), and three potato genotypes, namely CIP 381403.1, CIP 800212 BR 63-5 and 381379.9 (Kisoro). Muriate of potash (KCl) and urea were the K and N sources. The experimental design was a randomised complete block in a factorial arrangement, with three replications at each site. Hand-weeding and light hilling were done between 4 and 6 weeks after plant emergence. Crosses were made between Kisoro (female parent) and Rutuku (male parent) for production of hybrid TPS. These varieties were selected because of their high yield and compatibility. Both are popular varieties in Uganda. The other two genotypes, CIP 800212 and CIP 381403.1, were used for production of open-pollinated seed. Inflorescences and flowers on mother plants in the middle row of each experimental plot were counted every after 6 days, from the initial appearance of the flowers up to the end of bloom. Berries from the middle two rows of each plot were harvested separately and the corresponding number of mother plants recorded. Harvesting was done 5-6 weeks after full bloom, when the berries started to ripen. Berries from each plot were counted and weighed using a top-loading weighing scale (Ohaus GT-8000), and then ripened in polyethylene bags for six days at 27°C. Seeds were extracted onto a 0.5 mm sieve under running tap-water. They were then cleaned and dried at room temperature (27 + 3°C). A random sample of 5 g was taken from every seed lot and 100-seed weight determined using a mettler H-80 weighing scale. The seed was stored in paper envelopes in an incubator at 40 oC for 6 months, to allow seeds to break dormancy (Pallais, 1991). Data collected were analysed using the General Linear Models (GLM) Procedure (SAS, 1987) and least square means (LSMEANS) and probability difference matrices were used to determine levels of significance between the main treatment effects and the interactions. RESULTS Effect of N and K on flowering at Kalengyere. Flowering and berry set were generally low during the long rains (Table 1). Genotypes varied significantly (P≤0.05) in flowering with CIP 381403.1 producing more flowers and inflorescences per plant than CIP 800212. Variety Kisoro had significantly (P≤0.05) more flowers and inflorescences than the other two genotypes. Application of N and/or K, however, had no significant effect on flowering. Genotype x N x K interactions were also not significant (P≤0.05) during the season. During 1996 short rains, the flowering trend was reversed and was rather profuse compared to 1995 long rains (1995L). Kisoro variety, which had the highest flowering intensity during the previous season, produced less flowers and inflorescences than the other genotypes. Genotype CIP 800212 performed the best with 33.3 and 116.4 inflorescences and flowers per plant, respectively, compared to 1.7 and 3.2 inflorescences and flowers per plant, respectively, produced during 1995L. The performance of genotype CIP 381403.1 was intermediate (Table 1). Nitrogen application significantly (P≤0.05) increased flowering (flowers per plant). Joint application of N and K had no significant influence on the number of inflorescences during the season. However, the interaction of K x genotype was significant (P≤0.05) on this parameter. CIP 800212, flowered less with addition of K, while for CIP 381403.1, the converse was true (Fig. 1A). Kisoro, on the other hand, showed little response to K application. During 1999 short rains (1995S), CIP 381403.1 was not used because it was wiped out by bacterial wilt (Ralstonia solanacearum). Flowering was generally poorer than during (1996S) short rains. CIP 800212 produced 85% less flowers in the former than in the latter season (Table 1). Both inflorescence and flower production positively responded to N and K application. Nitrogen and K increased flowering by 43 and 26.5%, respectively. During 1999 L, mother plants hardly responded to N and K application and flowering was very poor in both CIP 800212 and Kisoro (Table 1). Effect of N and K on flowering at Namulonge. Flowering was generally low during 1996S, ranging from 0.6 and 14.9 flowers per plant, but genotypes differences was significant (Table 2). Genotype CIP 800212 produced the greatest number of flowers, followed by CIP 381403.1, which was trailed by Kisoro. Potassium application at 265.6 kg ha-1 significantly (P≤0.05) increased flowering, while N application had no significant effect on flowering during the season. During 1999S, flowering was moderate but greater than in 1996, and ranged from 15.7 to 26 flowers per plant (Table 2). There were significant (P≤0.05) genotypic flowering differences, with CIP 800212 producing 39.6% more flowers than Kisoro variety. Nitrogen did not influence flowering, but K significantly (P≤0.05) increased inflorescence produced per plant. There were no significant N x K interactions on these parameters. Flowering was lower during 1999L than in the previous season (1999S) (Table 2). For both CIP 800212 and Kisoro, flowering intensity was about half that of the previous season. However, inflorescence production was greater during the latter season (1999L). CIP 800212 produced significantly (P≤0.05) more inflorescences and flowers than Kisoro. Application of K significantly (P≤0.05) increased both parameters. Neither N application nor treatment interactions were significant on these parameters. Effect of N and K on berry set at Kalengyere. Berry production in genotypes CIP 800212 and CIP 381403.1 was poor during 1995L. Data for berry yield in these genotypes are, therefore, not presented here. The yield in Kisoro was, however, up to 30.7 berries per plant (Table 3). Nitrogen application at 240 kg ha-1 significantly reduced berry production per plant. Contrastingly, potassium application increased berry number and weight. Mother plants supplied with 265.6 kg K ha-1 had 26% more berries than the control plants. Application of 132.8 kg K ha-1 significantly (P≤0.05) increased berry weight. During 1996S, genotypes varied significantly in berry set (Table 3). Kisoro produced significantly (P≤0.05) more and heavier berries than the other genotypes. CIP 381403.1 produced the fewest berries, while CIP 800212 had the lightest berries. The number of berries produced per plant during the season was rather low and ranged from 8.5 to 12.2. Joint application of N and K significantly (P≤0.05) increased the number of berries per plant. Sole application of potassium at 265.6 kg ha-1 also significantly (P≤0.05) increased mean berry weight. However, sole N application had no significant influence on this parameter. Berry data for 1999 S and 1999L seasons do not include those for genotype CIP 381403.1 because it was destroyed by bacterial wilt. During the 1999S, berry production varied significantly (P≤0.05) between potato genotypes, with CIP 800212 producing 84.6% more berries than Kisoro (Table 3). Nitrogen application had no significant effect on berry production but K application significantly (P< 0.05) increased the number of berries per plant. Nitrogen x K interaction effects on this parameter were also significant (P< 0.05). Berry yield during the 1999L was very low and was not significantly influenced by the main effects of N and K (Table 3). However, N x genotype x K interactions were significant (P≤0.05) on this parameter (Fig. 1C and 1D). In genotype CIP 800212, K applied alone increased berry production up to 132.8 kg K ha-1, thereafter, berry production declined. Joint application of N and K at K rates higher than 132.8 kg ha-1 increased the number of berries per plant. In variety Kisoro, joint application of N and K, the latter at a rate less than 132.8 kg ha-1 depressed bery production, while higher K rates favoured berry production (Fig. 1D). Effect of N and K on berry set at Namulonge. During 1996S, there were significantly (P≤0.05) genotypic differences in berry set (Table 4). The number of berries per plant was 14.8 and 15.2 in potato genotypes CIP 800212 and CIP 381403.1, respectively. However, the two genotypes were not significantly different. Nitrogen application had no significant effect on the number of berries produced. However, K x genotype interactions were significant on this parameter (Fig. 1E). Potassium application rates less than 132.8 kg ha-1 depressed berry set in CIP 800212, while higher rates tended to increase the parameter in this genotype. On the other hand, the number of berries produced per plant in CIP 381403.1 increased linearly with K application (Fig. 1E). This was true irrespective of the N rate applied (Fig. 1H). Similarly, berry weight per plant and mean berry weight in CIP 381403.1 increased linearly with K application (Fig. 1F and 1G), respectively. The greatest total berry weight was 540 g, harvested from CIP 381403.1 plots (Table 4). In genotype CIP 800212, berry weight per plant declined with K application. During 1999S, only CIP 800212 was able to produce berries (Table 4). Berry yield was moderate and comparable to that obtained during 1996S. Berry weight per plant was not significantly influenced by N application. Potassium application, however, significantly (P≤0.05) increased both the number and weight of berries. Berry production during 1999L was lower than in the previous season and the response to N and K application was limited. There were no significant effects of either N or K on both the number and weight of berries during this season (Table 4). Berry weight per plot, however, significantly (P≤0.05) increased at K application rates higher than 132.8 kg ha-1. Effect of N and K application on true potato seed yield at Kalengyere. During 1995L, both number of seeds per berry and 100-seed weight were not significantly influenced by N and K application (Table 5). However, there were highly significant (P<0.01) K x N interactions on 100-seed weight (Fig. 2A). In the hybrid seed obtained from CIP 381379.9 (Kisoro) x Rutuku at Kalengyere, K increased seed weight at levels of N less than 120 kg ha-1 but depressed it at N levels ranging from 120 to 240 kg ha-1. Similarly, N increased TPS weight at K levels lower than 132.8 kg ha-1 and depressed it at higher K levels than these (Fig. 2A). Thus, there was a negative interaction between N and K on this parameter. During 1996S, the number of seeds per berry was generally less than that obtained during 1995L (Table 5) and was not significantly influenced by N and/or K application. The number of seeds per berry did not also differ significantly among the potato genotypes. There were, however, significant (P≤0.05) genotypic differences in 100-seed weight. Open-pollinated (OP) seeds from CIP 800212 and 'Kisoro x Rutuku' hybrid seeds were significantly (P≤0.05) heavier than OP seeds from CIP 381403.1 (Table 5). In addition, N and K application significantly (P≤0.05) increased TPS weight. During 1999S, CIP 800212 had significantly (P≤0.05) more seeds per berry than Kisoro and both N and K application significantly (P≤0.05) increased this parameter (Table 5). Nitrogen and K rates of 120 and 132.8 kg ha-1, respectively, were the most optimal in promoting seed production. An inverse relationship between 100-seed weight and number of seeds per berry was observed in genotype CIP 800212, which had more but lighter seeds than Kisoro. Furthermore, at K rate of 132.8 kg ha-1 mother plants produced the highest number of seeds per berry but the seeds were lighter than at the other K rates (Table 6). Nitrogen application had no significant effect on seed weight. There was also a significant (P≤0.05) N x K interaction on this parameter (Fig. 2B). Number of seeds per berry ranged between 53.7 and 71.2 and was not significantly affected by N application during 1999 long rains. On the contrary, potassium application significantly (P≤0.05) increased number of seeds during the season. True potato seed weight was quite high ranging from 96.8 to 113.3 mg in genotypes CIP 800212 and Kisoro, respectively. However, neither variety nor fertiliser main effects significantly influenced this parameter. Nitrogen x K interactions on seed weight were significant (P<0.01). Potassium alone increased seed weight, while joint N and K application depressed the parameter, indicating a negative interaction between the two nutrients (Fig. 2C). Effect of N and K application on true potato seed yield at Namulonge. There was evidence of genotypic differences in seed production per berry during the 1996S season (Table 6). The number of seeds produced per berry was 12.4% higher in CIP 381403.1 than in CIP 800212 during the season. However, neither N nor K significantly influenced this parameter. Similarly, 100-seed weight was significantly (P≤0.05) higher in CIP 381403.1 and was hardly affected by N and K application during the season (Table 6). Kisoro variety did not form berries at this location. During 1999S, neither K application nor genotypic effects had a significant influence on number of seeds and 100-seed weight. However, there was a significant but negative N x K interaction on 100-seed weight (Fig. 2D). Potassium rates higher than 132.8 kg ha-1 depressed seed weight, while lower rates increased the parameter. Number of seeds per berry was fairly high (68.1-97.4) during 1999L (Table 6). Nitrogen application did not influence this parameter during the season. However, a K rate of 132.8 kg ha-1 significantly (P≤0.05) increased seed number per berry. Seed weight was neither influenced by N nor K main treatment effects. However, there were significant (P<0.01) N x K interaction effects on TPS weight (Fig. 2E). Similar to 1999S, joint application of N and K depressed seed weight, especially, N rate of 120 kg ha-1. Nitrogen application rate of 240 kg ha-1 was not different from the control treatment (0 N kg ha-1)(Fig. 2E). DISCUSSION During 1995 long rains and 1999 short and long rains at Kalengyere, flowering was poor due to low soil moisture during the seasons. A dry spell set in before full bloom of the mother plants and this could have interfered with flower development (soil moisture measurements not taken). A typical response of plants to low soil moisture stress is the reduction in number of flowers and quantity of seed produced, without reducing the quality of seed. In this particular experiment, where soil moisture was inadequate, potato mother plants flowered early (about 40 days after planting compared to 50-60 days under normal circumstances) but produced very few flowers per plant. Flowering also varied significantly among genotypes and seasons. For example, Kisoro produced more flowers than other genotypes during the 1995L, but flowered the least during 1996S (Table 1). Seasonal effects are well reflected in the response of genotype CIP 800212, which produced a mean of 3.2 flowers per plant during 1995L compared to 116 flowers per plant during 1996S (Table 1). Seasonal influence on flowering is also indicated by the fact that flowering was better at Kalengyere than at Namulonge during 1996S, but much worse during 1999 short and long rains (Tables 1 and 2). Normally, flowering is favoured by cool weather (15-17 oC) (Marinus and Bodlaender, 1975). This is because most genotypes used were developed for the temperate climate that may only be simulated at high elevations in tropical areas (Sawyer, 1987). Although both genotypes, CIP 800212 and Kisoro, are fairly well adapted to warm and cool environments, only the latter flowered profusely under the warm environment at Namulonge. This is probably because most of the promising progenies that produce seed and ware potatoes from TPS are crosses whose male parents have been selected for the lowland tropics, and are associated with deficient flowering under short-day conditions (Pallais et al., 1984). Genotypic differences in growth and development of potatoes are well documented (Marinus and Bodlaender, 1975) and differences in flowering among potato genotypes have been reported (Sadik, 1984; Upadhya et al., 1984; Kanzikwera et al., 1997). These differences are a reflection of the influence of factors such as temperature, photoperiod, available moisture and nutrients in the soil. Varieties in the same maturity class have been shown to have different responses to temperature in flowering and plant growth habits (Marinus and Bodlaender, 1975). It is, therefore, not surprising that genotypic differences in flowering were observed in this study. Reduced flowering among the genotypes may also be attributed to genetic influence. Eguchi (cited by Pallais et al., 1984) observed that flower differentiation takes place during tuber sprouting. Competition for assimilates by other developing organs could, therefore, limit floral development. Recent advances in plant molecular biology have revealed floral meristem identity genes in Arabidopsis (Yanofsky, 1995). Furthermore, early-acting genes that promote formation of floral meristem and later-acting genes that determine the fate of floral organ primordia have been identified. This information will be useful in understanding the flowering process as well as the survival of flowers in potato. Generally, there was either no or low influence of N application on flowering
of TPS mother plants. This was probably due to the relatively lower N rates
(highest rate = 240 kg ha-1) used in this study, compared to those
used by other workers (Pallais et al., 1987). Moreover, tubers were not
pruned in order to reduce underground competition, and this may have contributed
to the poor response of mother plants to N application. Pallais et al.(1987),
for example, used N rate of 1000 kg ha-1 applied twice, half at planting
and the other half at 30 days after planting, in addition to tuber removal.
Increased flowering in mother plants through split applications of N have been
reported (Pallais et al., 1987; Maingi et al., 1994). The split
application effect was, however, not investigated in this study. Also, since
flowering in potatoes coincides with the phase of rapid tuber enlargement, and
increased potato yields depend on assimilate mobilisation from the aerial structures
to the developing tubers, reduced flowering, berry set and seed development
may have been caused by this phenomenon (Pallais et al., 1987). The effect of K on TPS weight has not been reported although its influence on fruit yield is well known. Albregts et al. (1991) reported a positive linear fruit weight response to K in strawberry (Fragacia x ananassa Duch.). They found that leaf K concentrations correlated well with the K rates from 56 to 224 kg ha-1. In the present study, K applied alone increased seed weight and favoured K accumulation in the berries. The increase in seed weight due to K application may be related to its role in protein synthesis as well as K's own weight. Potassium is required for protein synthesis in higher plants and is thought to be involved in the translation and binding of tRNA in the ribosomes (Marschner, 1990). In K deficient and K sufficient tobacco plants, total 15N taken up and incorporated into protein N was 11 and 32%, respectively (Marschner, 1990). In conclusion, this study confirms potato varietal differences in flowering, berry and seed set, and response to N and K application. Potassium application increases flowering, berry set and tuber yield, but this was greatly influenced by location and season. Potato mother plants respond poorly to N and K application in flowering, berry set and TPS yield. This is attributed to competition between floral parts and tubers for growth resources and to a negative interaction between the two nutrients. Joint N and K application may depress TPS weight and dry matter production due to both direct and indirect negative influence of these nutrients on the uptake of other nutrients especially, Ca and Mg. ACKNOWLEDGEMENTS This work was done with the support of the National Agricultural Research Organisation (NARO) and United States Agency for International Development (USAID)-Washington through a collaborative research grant to Makerere University and Fort Valley State University, Georgia, USA. REFERENCES
TABLE 1. Effect of nitrogen and potassium fertilisation on flowering of true potato seed mother plants at Kalengyere Research Station
Values followed by the same letter or not followed by a letter
are not significantly different at P≤0.05. *=significant at P≤0.05, NS
= not significant at P≤0.05
TABLE 2. Effect of nitrogen and potassium fertilisation on flowering of true potato seed mother plants at Namulonge Research Institute
Values followed by the same letter or not followed by a letter
are not significantly different at P<0.05
NS = not significant at P<0.05, IPP = Number of inflorescences per plant, FPP = number of flowers per plant TABLE 3. Effect of nitrogen and potassium application on berry production in true potato seed mother plants at Kalengyere Research Station
Values followed by the same letter or not followed by a letter
are not significantly different at P≤0.05. *=significant
at P≤0.05, NS = not =significant at P≤0.01
BPP = Number of berrnt, MBWT = Mean berry weight (g) TABLE 4. Effect of nitrogen and potassium application on berry production in true potato seed mother plants at Namulonge Research Institute
Values followed by the same letter or not followed by a letter
are not significantly different at P<0.05. *= significant at P<0.05, **=significant
at P<0.01, ***=significant at P<0.001, NS = not significant at P≤0.05;
BPP = Number of berries per plant, MBWT= mean berry weight (g), BWTPP = berry
weight per plot (g)
TABLE 5. Effect of nitrogen and potassium application on True Potato Seed (TPS) yield at Kalengyere Research Station
Values followed by the same letter or not followed by a letter
are not significantly different at P<0.05. NS = not significant at P<0.05
**=significant at P<0.01, SPB =number of seeds per berry, TPSWT =weight of 100 seeds (mg) TABLE 6. Effect of nitrogen and potassium application on True Potato Seed (TPS) yield at Namulonge Research Institute
Values followed by the same letter or not followed by a letter
are not significantly different at P<0.05. NS = not significant at P<0.05,
**=significant at P<0.01
SPB = Number of seeds per berry,TPSWT = weight of 100 seeds (mg) The following images related to this document are available:Line drawing images[cs01041f.gif] [cs01041b.gif] [cs01041k.gif] [cs01041e.gif] [cs01041c.gif] [cs01041m.gif] [cs01041l.gif] [cs01041g.gif] [cs01041d.gif] [cs01041j.gif] [cs01041h.gif] [cs01041i.gif] [cs01041a.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}