 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 9, No. 1, March 2001, pp. 127-146
C. R. Kanzikwera, J. S. Tenywa1, D. S. O Osiru, E. Adipala and A. S. Bhagsari2
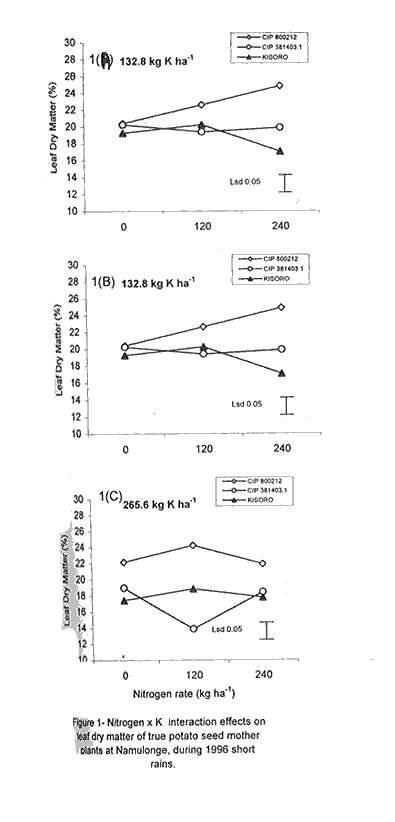
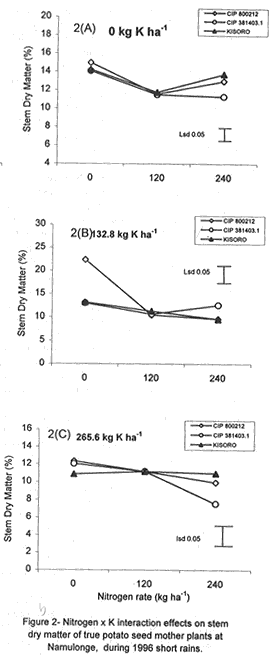
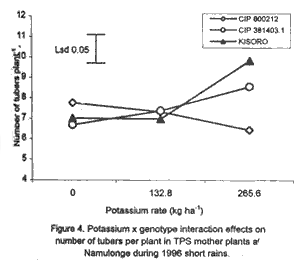
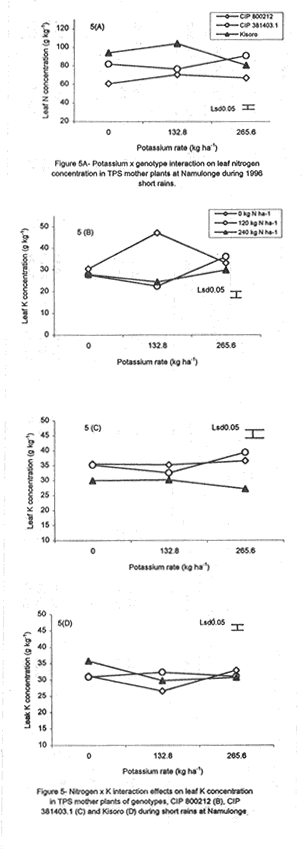
Code Number: CS01042 ABSTRACT Field experiments were conducted at Namulonge Agricultural and Animal Production Research Institute and Kalengyere Research Station in Uganda, during 1995-1996 and 1999, to assess the effect of N and K on dry matter yield and nutrient partioning in true potato (Solanum tuberosum) seed (TPS) mother plants. Three N (0, 120, 240 kg ha-1) and K (0, 132.8 and 265.6 kg ha-1) rates were applied to mother plants of three potato genotypes, CIP 800212, CIP 381379.9 (Kisoro) and CIP 381403.1. Potassium application significantly (P≤0.05) depressed shoot dry matter yield in all the genotypes. Nitrogen application, however, had no significant effect on shoot dry matter yield although N x genotype interactions were significant (P≤0.05) on the parameter. Fresh tuber yield ranged from 21.0 to 37.5 t ha-1, and was significantly (P≤0.05) increased by both N and K application. Leaf N concentration varied significantly (P≤0.05) among genotypes and K rates higher than 132.8 kg ha-1 increased this parameter in potato genotype CIP 381403. High N and K rates also increased stem N concentration in this genotype. Nitrogen application significantly (P≤0.05) increased foliar Ca concentration. In genotype CIP 800212, K application depressed foliar Mg concentration in the absence of applied N. Leaf Mg concentration declined at K application rate less than 132.8 kg ha-1. Potassium significantly (P≤0.05) increased leaf P concentration, while N depressed this parameter. Stem K concentrations varied significantly (P≤0.05) among the potato genotypes. Nitrogen application increased stem K concentration, while K reduced this parameter. There was significant N x K interaction on stem Mg concentration. Both N and K significantly (P≤ 0.05) increased berry P and Ca. Nitrogen and K were found to have a negative interaction on Ca, Mg, K, N and P concentrations in the leaves, stems and berries of TPS mother plants. Key Words: Nutrient antagonism, potato genotypes, Solanum tuberosum, tuber yield RÉSUMÉ Des essais en champs ont été conduits à l'Institut de Recherche de production animale et agricole à Namulonge et à la station de recherche de Kalengyere en 19995, 1996 et 1999 pour évaluer l'effet de l'N et du K sur la floraison, l'établissement des baies et sur le rendement des TPS et leur influence sur les éléments nutritifs. Trois doses d'Azote (0, 120, 240 kg ha-1) et de K (0, 132.8 ET 265.6 kg ha-1 ) ont été appliquées sur les plantes- mères des vraies semences de 3 génotypes de pomme de terre, CIP 800212, CIP 381379.9 (Kisoro) et CIP 381403.1. L'application du potassium a réduit significativement (P≤0.05) la matière sèche de tous les génotypes. Cependant l'application de l'Azote n'a pas eu d'effet significatif sur la matière sèche des feuilles bien que les interactions entre l'N et le génotype pour ce paramètre était significative (P≤0.05). Le rendement des tubercules frais variait entre 21 et 37.5 t ha-1 et était significativement favorisé par l'application de l' N et du K. La concentration en N de la feuille variait significativement (P≤0.005) parmi les génotypes et la dose élevée de K de 132.8 kg ha-1 a favorisé ce paramètre chez le génotype CIP 381403. Des taux élevés d' N et de K ont augmenté la concentration de l'N dans les tiges chez le même génotype. L'N a significativement augmenté la concentration foliaire du Ca. L'application du K a réduit chez le génotype CIP 800212, la concentration du Mg en l'absence de l'N. La concentration du Mg des feuilles chez les plantes-mères a baissé suite à une application de K inferieure à 132.8 kg ha-1. Le potassium a augmenté significativement (P≤0.05) la concentration du phosphore dans les feuilles, alors que l'N a réduit cet élément. La concentration du K dans les tiges a significativement (P≤0.05) varié entre les génotypes. L'application de l'N a influencé l'augmentation du P foliaire alors que le K a réduit ce facteur. Il y avait une interaction entre N et K sur la concentration du Mg de la tige. Ensemble l' Azote et le Potassium ont augmenté significativement (P≤0.05) le Ca et le P des baies. L'N et le K ont eu une interaction négative sur les concentrations du Ca, Mg, K, N et P des feuilles, tiges et des baies des plantes-meres de TPS. Mots Clés: Nutrient antagonism, potato génotypes, Solanum tuberosum, tuber yield INTRODUCTION The key to growth and development in a potato (Solanum tuberosum) crop, and to its ultimate yield, lies in the balance between the conflicting resource demands of mainly two sinks, foliage and growing tubers (Hay and Walker, 1989; Walter et al., 1990). Potato tuber yield is directly dependent on the supply of nitrogen, potassium and phosphorus, though excessive supply of N may substantially delay leaf senescence leading to enhanced leaf area duration and increased tuber yield (MacKerron and Heilbronn, 1985). However, Dyson and Watson (1971) and Sattelmacher and Marschner (1979) showed that N application also delays tuber initiation and bulking. Furthermore, excessive N supply increases flower production and pollen germination and promotes production of vigorous and high quality TPS (Pallail et al., 1984; Pallais et al., 1987; Pallais, 1991; Maingi et al., 1994). Potassium plays major physiological and biochemical roles in plant growth (Beringer et al., 1990). It is very essential in protein synthesis and activation of enzymes, besides enhancing transport of photosynthate from leaves to other plant parts. It is also very useful in osmo-regulatory mechanisms in plants. In earlier studies (Kanzikwera et al., 1997), TPS mother plants responded poorly to K application in terms of flowering, berry set and TPS production. This poor response was attributed to competition between tubers and aerial plant parts for limited assimilates, and to low N and reduced uptake of Mg and Ca due to K-Mg and K-Ca antagonisms. This study was conducted to determine the effect of N and K application on N, K and other mineral nutrients; as well as leaf, stem, berry and tuber dry matter yield of TPS mother plants. The effects of these nutrients on seed weight and seedling emergence are described in detail by Kanzikwera et al. (2000). In this volume we have also described the effects of these nutrients on flowering and berry set of true potato seed mother plants (see Kanzikwera et al., 2001). MATERIALS AND METHODS The experiment was conducted during four cropping seasons (1995-1996 and 1999) at Kalengyere Research Station (KRS) and three seasons (two in 1996 and one in 1999) at Namulonge Agricultural and Animal Production Research Institute (NAARI). Kalengyere lies (1° 45'S, 29o 15'E) in southwestern Uganda, at an altitude of 2,450 metres above sea level, with average annual rainfall of 1,250 mm, biannually distributed. The first season has long rains, while the second has a shorter duration. The area is relatively cooler than Namulonge, with 17 °C average daily temperatures compared to 21°C at Namulonge. The soils are volcanic (Andosols) and fertile with patches of lava-ash. The soil characteristics of the two sites in 1995 are shown in Table 1. Treatments consisted of three levels of K (0, 132.8 and 265.6 kg ha-1) and N (0, 120 and 240 kg ha-1), and three potato genotypes, namely CIP 381403.1, CIP 800212 (BR 63-5), and 381379.9 (Kisoro). Muriate of potash (KCl) and urea were the K and N sources, respectively. The experiment, which was factorial in nature, was laid out in a randomized complete block design with three replications at each site. Hand-weeding and light hilling were done between 4 and 6 weeks after plant emergence. Leaf samples consisting of the most recently matured leaf petiole and blade, and samples of plant shoots, berries and tubers, were collected from four plants in the middle row of each plot. This was done when the first cluster of berries was forming (60 days after planting). The samples were cleaned using a detergent solution, then tap-water and finally distilled water. They were oven-dried at 70 °C for 48 hr to a constant weight. Subsequently, they were allowed to cool at room temperature in a desiccator and weighed immediately. Tuber, stem, berry and leaf dry matter contents were determined gravimetrically. The samples were ground to pass through a 1.0 mm sieve, digested for two hours in concentrated sulphuric acid, and then analysed for total N, K and Ca and Mg using the procedures outlined by Okalebo et al. (1993). Data collected were analysed using the General Linear Models (GLM) procedure (SAS, 1987) and least square means (LSMEANS) and probability difference matrices were used to determine levels of significance between the main treatment effects and the interactions. Relationships between fertiliser treatments and TPS-related parameters were determined using regression and correlation analyses. RESULTS Dry matter partitioning and fresh tuber yield response to N and K application. At Kalengyere, shoot dry matter (SHDM) ranged from 12.1 to 14% and was significantly (P≤0.05) higher in CIP 800212 and Kisoro than in CIP 381403.1 (Table 2). Potassium application depressed shoot dry matter in all genotypes at this site. Nitrogen application, however, had no significant effect on yield, though N x genotype interaction effects on the parameter were significant (P≤0.05). There was evidence of a seasonal effect on shoot dry matter. Both N and K application significantly (P≤0.05) depressed the parameter. In addition, K application depressed tuber dry matter (TDM) yield during the season. Fresh tuber yield (FTY) was significantly (P≤0.05) increased by joint application of N and K but not by K alone (Table 3). The number of tubers per plant (TPP) increased from 6.8 in the control mother plants to 8.6 in the mother plants supplied with 265.6 kg K ha-1. Potassium application rates greater than 132.8 K ha-1, however, did not cause a significant effect. Nitrogen and K increased tuber yield per unit area by 21.3 and 41.2%, respectively, over the control (0 N + 0 K kg ha-1). Similarly, tuber weight increased due to N and K application. Nitrogen application rate of 240 kg ha-1 increased tuber weight by 23% over the control. This compared well with the 25% increase when 265.6 kg K ha-1 was applied to the mother plants. Genotype CIP 381403.1 produced heavier tubers than CIP 800212 and Kisoro during 1995 long rains, but this changed during 1999 short rains and long rains (1995L) when Kisoro outweighed CIP 800212. Genotype CIP 381403.1 had the highest fresh tuber yield during 1996 short rains, while yields of Kisoro and CIP 800212 were not significantly different (Table 2). Fresh tuber yield per unit area also increased with K application, though K rates greater than 132.8 kg ha-1 caused no significant effect. Number of tubers produced per plant significantly (P≤0.05) increased with N and K application. Tuber dry matter was influenced by N rate x genotype interactions (P≤0.01) at Kalengyere. Potato genotypes varied in response to N application in terms of tuber dry matter yield, though N generally increased the parameter (Table 2). Nitrogen application and tuber dry matter were positively and linearly related in the case of genotype CIP 800212. In genotype CIP 381403.1, however, N rates less than 150 kg ha-1 depressed tuber dry matter. On the contrary, in Kisoro variety, tuber dry matter increased at K rates lower than 132.8 kg ha-1 and declined at higher K application rates. At Namulonge, where more detailed dry matter partitioning was studied, leaf dry matter (LDM) content varied significantly among genotypes and was highly influenced by N and K application (Fig. 1A). Potassium x N x genotype interactions were also highly significant (P≤0.001). For this reason, main treatment effects on leaf dry matter are not presented here. Without K application (Fig. 1A), leaf dry matter in CIP 800212 potato mother plants declined whenever N was applied. In CIP 381403.1 mother plants, however, leaf dry matter increased at low levels of N up to 120 kg ha-1, and gradually declined at rates higher than this (120 kg N ha-1). There was less response to N in Kisoro mother plant compared with other genotypes. At the K rate of 132.8 kg ha-1, leaf dry matter in CIP 800212 was positively and linearly related to N application (Fig. 1B). In CIP 381403.1 mother plants, however, leaf dry matter was depressed by N application. In Kisoro variety, leaf dry matter increased with N application up to a peak (120 kg ha-1) and then declined. At the highest K application rate (265.6 kg ha-1), leaf dry matter in CIP 800212 and Kisoro increased with N application up to a maximum and declined thereafter. In CIP 381403.1, however, N depressed leaf dry matter (Fig. 1C). At the same site, stem dry matter content (SDM) was significantly (P≤0.05) higher in CIP 800212 than in genotypes CIP 381403.1 and Kisoro, both of which did not differ significantly. Both N and K application depressed shoot dry matter. The parameter was also significant (P≤0.001) influenced by N x K x genotype interaction (Fig. 2A). In the absence of applied K (Fig. 2A), shoot dry matter declined with N application in all genotypes. The decline, however, was more pronounced in CIP 381403.1 than in the other genotypes. At the K rate of 132.8 kg ha-1 (Fig. 2B), there was a distinct drop in shoot dry matter in CIP 800212 mother plants as N rates increased. In the other genotypes, however, shoot dry matter declined with N application. At 265.6 kg K ha-1 (Fig. 2C), shoot dry matter declined linearly with N application in genotype CIP 800212. CIP 381403.1 response was similar but the decline was steeper for the latter genotype. There was little response to N in Kisoro's shoot dry matter. Berry dry matter content (BDM) was influenced by N rate x genotype interaction (Fig. 3A). In both CIP 800212 and CIP 381403.1 genotypes, N application increased berry dry matter, though the increment gradually diminished with a higher N application rate. CIP 381403.1 berry dry matter responded better to N as reflected by the higher rate of change in berry dry matter with N application (Fig. 3A). Both N and K influenced tuber dry matter content significantly (P≤0.05) (Fig. 3B and Fig. 3C). Genotypes CIP 381403.1 and Kisoro responded similarly to N application. In fact, the response graphs of both genotypes overlap. In both genotypes, TDM increased but the increments diminished at a higher N application rate (Fig. 3B). Tuber dry matter yield of CIP 800212 was little influenced by N application. Neither N nor K application impacted the number of tubers per plant or tuber yield per unit area significantly (P≤0.05) (Table 3). However, there were significant K x genotype interaction effects on number of tubers per plant (Fig. 4), although this parameter declined linearly with K application in CIP 800212. In CIP 381403.1, however, K application increased the parameter linearly. In Kisoro variety, a high rate of K application favoured tuber dry matter. Fresh tuber yield ranged from 8.4 to 9.3 t ha-1, but was not significantly (P≤0.05) affected by any of the treatments (Table 3). Similarly, mean tuber weight was hardly affected by treatment factors. Foliar nutrient concentration. Nitrogen was the nutrient with the highest concentration (81.03 mg kg-1) in the leaves (Table 5). This was followed by K (31.9 mg kg-1), Ca (13.97 mg kg-1), phosphorus (3.03 mg kg-1), Mg (2.3 mg kg-1) and, lastly by Na (0.1 mg kg-1). Generally, N application had a significant (P≤0.05) influence on K, Ca and Mg concentrations in TPS mother plants; there were also significant K x N interactions on leaf K and Ca concentrations. Sole K application, however, had no significant (P≤0.05) effect on the quantitative nutrient status in the leaves, although K x genotype interaction effect on leaf N was significant at P≤0.05 (Table 5). There were significant (P≤0.01) N x genotype and K x N x genotype interaction (P≤0.001) on leaf K concentration. Leaf N concentration was the lowest in CIP 800212 genotype (66.3 mg kg-1) (Table 5), and highest in Kisoro (93,3 mg kg-1). CIP 381403.1 had intermediate leaf N concentration (83.4 mg kg-1). The effect of K application on leaf N concentration varied among genotypes (Fig. 5A). In genotypes CIP 800212 and Kisoro, K less than 132.8 kg ha-1 increased leaf N, but a K rate greater than this led to a decrease in leaf N. On the contrary, in CIP 381403.1, K rates higher than 132.8 kg ha-1 increased leaf N concentrations (Fig. 5A). With regard to mother plants foliar P concentration in TPS, genotypes CIP 800212 and Kisoro had similar but significantly (P≤0.05) higher values than CIP 381403.1 (Table 5). Neither N nor K application influenced foliar P concentration significantly (P≤0.05). Similarly, foliar K concentration varied significantly (P≤0.05) among the genotypes, as did K x N x genotype interactions (Table 5). Generally, there was a negative interaction caused by joint N and K application on foliar K, though the variation was inconsistent among the genotypes (Figs. Fig. 5B, 5C and 5D). Figures 5B and 5C show the influence of N and K on foliar K concentration in genotype CIP 800212. Without N application, foliar K increased with K application up to a peak (132.8 kg ha-1), and then declined. This scenario completely reversed with N application. The effect of N and K application on foliar K concentration in Kisoro variety, however, contrasted with genotypes in that, joint N and K application increased foliar K (Fig. 5D). Foliar Ca concentrations were not significantly different among the genotypes
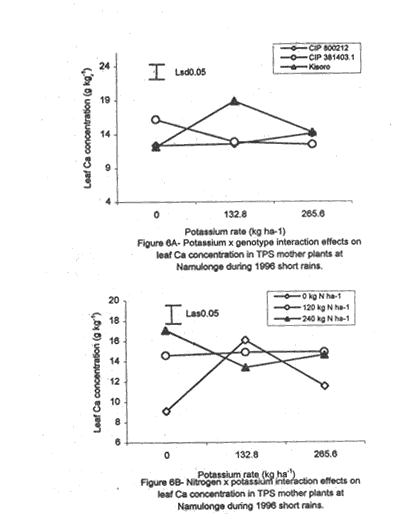
and were not affected by K as a main treatment (Table 5). However, N application
significantly (P≤0.05) increased foliar Ca concentration. There were also
significant (P≤0.05) K x genotype and K x N interactions effects on this
parameter (Fig. 6A). The influence of K on foliar Ca concentration again varied
among the genotypes. In Kisoro, leaf Ca increased with K application up to a
peak (19 mg kg-1), and then declined at K rates greater than 132.8 kg ha-1.
In contrast, leaf Ca in CIP 381403.1 declined with K application, and was the
exact opposite of foliar Ca in CIP 800212 mother plants, which increased linearly
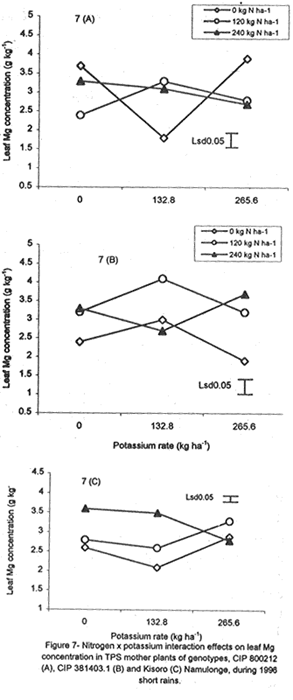
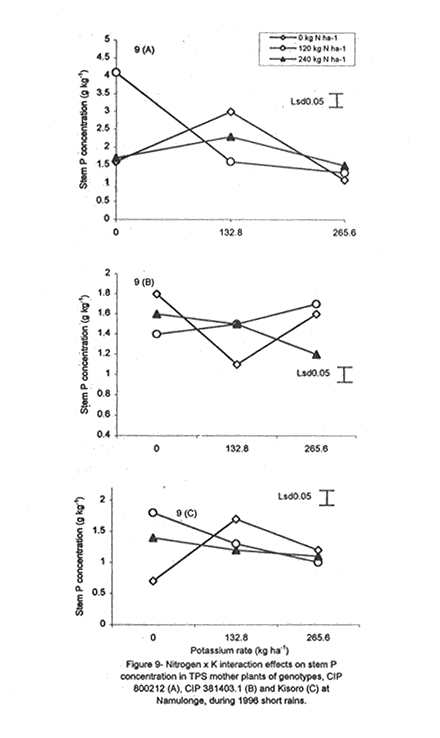
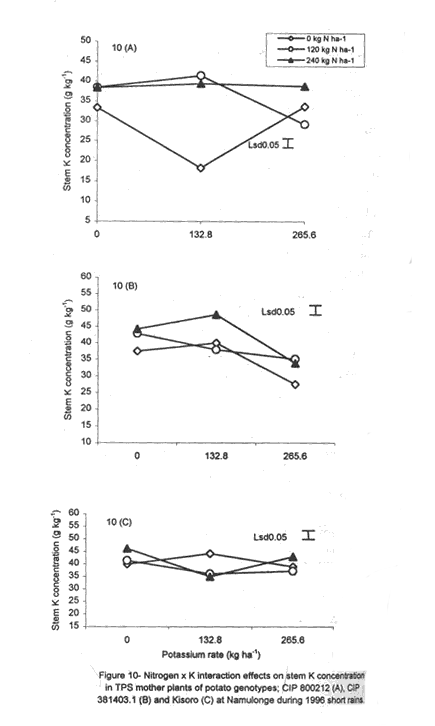
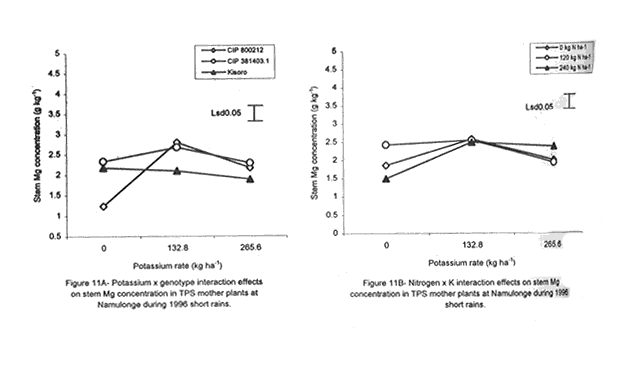
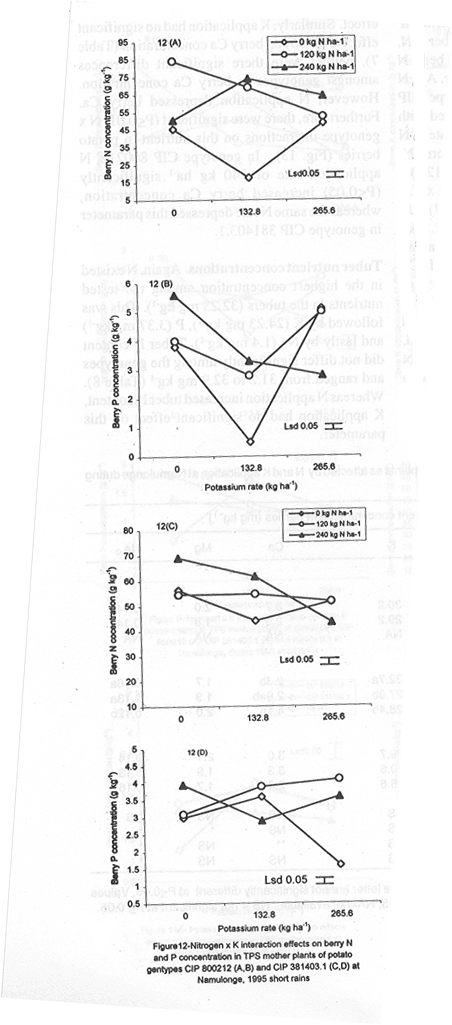
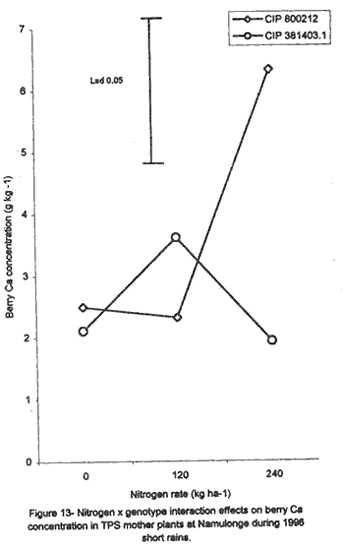
with K application (Fig. 6A). Leaf Mg did not vary significantly (P≤0.05) among the potato genotypes (Table 5). Nitrogen application increased foliar Mg, while potassium had no significant (P≤0.05) effect. There were significant (P≤0.05) K x N x genotype interaction effects on this parameter. The effect of N and K on foliar Mg concentration varied among genotypes and is shown in Figure 7A. In CIP 800212, K application generally depressed foliar Mg concentration in potato not fed with N. In mother plants supplied with 120 kg N ha-1, Mg peaked at K rate of 132.8 kg ha-1 and, thereafter, declined. In the case of mother plants fed with a higher N rate (240 kg ha-1), Mg concentration declined linearly with K application. In contrast, K application increased foliar Mg concentration in genotype CIP 381403.1 up to a peak (132.8 kg ha-1) after which there was a decline in this parameter (Fig. 7B). This was particularly so in mother plants that fed with none or 120 kg N ha-1. Leaf Mg concentrations in cases fed with 240 kg ha-1 declined, at K application rates less than 132.8 kg ha-1. However, higher K rates, increased Mg concentrations. Similar responses were obtained with Kisoro variety. Generally, however, mother plants that received N had higher foliar Mg concentrations than the controls. Sodium concentration in the leaves was stable at 0.1 g kg-1 and was not significantly affected by the treatments (Table 5). Stem nutrient concentrations. Stem N concentration did not differ significantly among potato genotypes; neither did N nor K application cause a significant effect (Table 6). However, there were significant K x genotype (P≤0.001), K x N (P≤0.01), N x genutype (P≤0.05) and K x N x genotype (P≤0.01) interactions on this parameter. In CIP 800212 (Fig. 8A), N and K application caused inverse effects on stem N concentrations. The effect on CIP 381403.1 was different (Fig. 8B); N and K application positively increased stem N concentrations in this genotype. In Kisoro variety, K rates higher than 132.8 kg ha-1 accompanied by a N rate of 120 kg ha-1 increased stem N (Fig. 8C). With respect to stem P concentrations, genotype CIP 800212 had significantly higher (P≤0.05) values than the other two genotypes (Table 6). Sole potassium application reduced stem P concentrations significantly (P≤0.05). Applica-tion of N, however, had no significant effect on this parameter. There were significant K x N (P≤0.05) and K x N x genotype (P≤0.01) interactions on P concentrations. In CIP 800212, low rates of K (<132.8 kg ha-1) increased leaf P concentration, while N application rate of 120 kg ha-1 and K rate of 265.6 kg ha-1 depressed this parameter (Fig. 9A). In CIP 381403.1, stem P concentration increased linearly with K at a fixed N application of 120 kg ha-1. Conversely, leaf P declined linearly with K application at a N rate of 240 kg ha-1 (Fig. 9B). In Kisoro, N and K rates greater than 150 kg ha-1 reduced stem P, while lower rates than these increased the parameter (Fig. 9C). Stem K concentrations varied significantly (P≤0.05) among potato genotypes, with CIP 381403.1 and Kisoro having higher stem K than CIP 800212 (Table 6). Nitrogen application increased stem K concentration, while K application depressed this parameter. Nitrogen x K x genotype interactions were also significant (P≤0.05). There was a negative interaction between N and K in CIP 800212 (Fig. 10A) and CIP 381403.1, whereby high rates of K and N reduced stem K concentration (Fig. 10B). In Kisoro, N application depressed stem K concentration, while K application increased it (Fig. 10C). At high K rates (>132.8 kg ha-1), however, stem K concentration was reduced. TPS mother plant content of Ca in stems was significantly (P≤0.05) genotypically influenced, being higher in Kisoro than in the other genotypes (Table 6). Both N and K application had no significant effect on this parameter. Magnesium concentration in the stems was strongly influenced by K x genotype (P≤0.001) and K x N (P≤0.05) interactions (Table 6). In all genotypes, K application rates below 132.8 kg ha-1 increased stem Mg; yet, higher K rates decreased it (Fig. 11A). The increase in Mg concentration was highest in CIP 800212 and the least in Kisoro. Generally, there was a negative interaction between N and K on stem Mg. In CIP 800212, stem Mg concentrations increased at higher rates of K application (>132.8 kg ha-1), but the converse was true in the other two genotypes (Fig. 11B). On the other hand, stem Na concentration was constant and not significantly (P≤0.05) affected by the treatments. Potassium was the nutrient in highest concentration (37.83 mg kg-1) in stems. Berry nutrient concentrations. Among the nutrients evaluated, N existed in the greatest concentrations (mean = 55.17 mg kg-1) in potato berries across genotypes. Again, Na concentration ranked the lowest (Table 7). CIP 800212 and CIP 381403.1 did not differ significantly in berry N. Nitrogen application generally increased berry N content, while K depressed the parameter. At a N rate of 120 kg ha-1, berry N in genotype CIP 800212 was negatively and linearly related with K application (Fig. 12A). At the higher rate of N (240 kg ha-1), K application depressed berry N. This was also true in CIP 381403.1 (Fig. 12B). Berry P was significantly affected by N x K x genotype interactions (P≤0.001) (Table 7). In CIP 800212, a K application rate of 132.8 kg ha-1 depressed berry P, while higher K rates along with N application increased berry P (Fig. 12C). In CIP 381403.1, however, joint application of high N and K depressed berry P (Fig. 12D). Application of N at a rate of 120 kg ha-1 increased berry P markedly as K application was increased. The concentration of K in potato berries did not differ significantly (P≤0.05) among the potato genotypes (Table 7). Moreover, berry K concentrations were depressed by N application, but K application had no significant (P≤0.05) effect. Similarly, K application had no significant effect (P≤0.05) on berry Ca concentration (Table 7), neither were there significant differences amongst genotypes in berry Ca concentration. However, N application depressed berry Ca. Furthermore, there were significant (P≤0.01) N x genotype interactions on this nutrient in potato berries (Fig. 13). In genotype CIP 800212, N application rate of 240 kg ha-1 significantly (P≤0.05) increased berry Ca concentration, whereas the same N rate depressed this parameter in genotype CIP 381403.1. Tuber nutrient concentrations. Again, N existed in the highest concentration among the tested nutrients in the tubers (32.23 mg kg-1). This was followed by K (24.23 mg kg-1), P (3.37 mg kg-1) and lastly by Na (1.4 mg kg-1). Tuber N content did not differ significantly among the genotypes and ranged from 31.2 to 32.9 mg kg-1 (Table 8). Whereas N application increased tuber N content, K application had no significant effect on this parameter. Tuber P concentrations differed significantly (P≤0.05) among the potato genotypes (Table 8) being highest in CIP 800212 (3.6 mg kg-1) and lowest in CIP 381403.1 (3.1 mg kg-1). However, neither N nor K had a significant effect on P concentration in the tubers. Tuber K content was significantly (P≤0.05) higher in CIP 800212 and Kisoro than in CIP 381403. There were consistent effects of potassium application on tuber K concentrations. The concentrations of Ca and Mg in the tubers ranged from 1.1 to 1.4 mg kg-1, but were not significantly affected (P≤0.05) by the treatments (Table 8). DISCUSSION Joint application of K and N generally lowered dry matter content in potato mother plants. This parameter was also found to be influenced by N, K and genotype interactions. Potassium application generally depressed tuber dry matter yields (Tables 2 and 3). High K fertilisation, especially, in the form of muriate of potash (KCl), has been reported to depress dry matter content of potato tubers (Burton, 1966). This yield decline could be related to the repressive effect of K on starch synthase. In this study potassium was found to be the second highest contained nutrient in potato tubers (Table 8) and this could have led to reduced tuber dry matter production. The depressive effect of applied N on tuber dry matter also conforms to the observations of Cole (1975), whereby tuber dry matter (per cent of fresh weight, commonly expressed in terms of specific gravity) fell with increased N application. Dry matter content may be related to the state of juvenility (or tenderness) and age of the mother plants (Williams et al., 1995). Mature plants usually have higher dry matter than young ones. Nitrogen and K application could have induced delayed senescence of mother plants, leading to a decline in dry matter of leaves and stems (Burns, 1992). This may also be associated with the influence of these nutrients on cytokinin and other phytohormonal activities which have direct influence on plant growth and dry matter accumulation. Observations in the field conform to this view because mother plants that received high rates of N and K took longer to senesce, than the control plants. Among the potato genotypes tested, CIP 381403.1 showed more positive response to K in berry dry matter yield than the other genotypes. Potassium concentration in the leaves was also fairly high in this genotype (Table 5). It is possible that in CIP 800212 and Kisoro (early-maturing genotypes), N and K were translocated from the aerial plant parts to the tubers (dominant sinks) early in the growing season. In early maturing potato varieties, tuber bulking also starts early and coincides with the time of flower development and berry set (Hay and Walker, 1989). In these genotypes, therefore, less K may have been channelled into flower production. This view is supported by the significantly higher tuber K concentration found in these early maturing genotypes (Table 8) compared to that of CIP 381403.1 (late maturing). Translocation of nutrients during early growth stages, from leaves and stems to developing organs, has been reported elsewhere (Kirkby and Mengel, 1967). Differences in K concentrations observed in the potato genotypes may be related to differences in K uptake capacity resulting from plant morphological and structural differences. Cassman et al. (1989) and Brouder and Cassman (1990) reported differential K uptake in cotton cultivars. They related this to differences in root length development, root mean diameter or both. Racher et al. (1999) also observed similar results with 23 bean genotypes growing under low P supply. Lack of response to N and the low response to K in berry dry matter yield in this experiment, however, is mainly attributed to competition between the tubers and the above ground plant parts. This is reflected in the high tuber yields obtained at Kalengyere during 1995 long rains, 1996 short rains and 1999 long rains (Tables 2 and 4), and a high rate of nutrient partitioning into these plant parts (Table 8). The negative interaction between N and K seems to be related to the influence of these two nutrients on the concentration of other mineral nutrients in the mother plants (Bester and Maree, 1990). Potassium application in disproportionate quantities is associated with decline in Mg and Ca concentration in the mother plants as shown in Figures 6 and 7. High rates of K also reduced berry N and K (Fig. 9). Nitrogen application, on the other hand, increased berry P, Ca, Mg and N. Similar negative interactions between N and K have been reported in maize by Karlen and Whitney (1980) and Karlen et al. (1987). According to these researchers, excess K accumulation may interfere with N uptake and reduce grain yield. The concentration of nutrients in leaves of potato mother plants was basically similar in all the potato varieties (Table 8). However, genotypes CIP 381403.1 and Kisoro accumulated higher N than did CIP 800212. On the other hand, CIP 381403.1 had the lowest leaf P concentration. Excessive accumulation of K was reported to lower P levels in maize due to its negative interaction on N (Karlen et al., 1987). Application of N increased leaf Mg and Ca, but reduced leaf K concentration in the present study. This further confirms the observation made above that the negative K and N interactions on potato mother plants is a reflection of their influence on other mineral nutrients. Potassium was the most abundant nutrient in potato stems (Table 6). This is probably due to its key functions in phloem transport. Marschner (1990) reported higher K+ concentrations in stems of higher plants than any other nutrient and related this to its role in the sucrose phloem loading mechanism. Potato berries accumulated nutrients (Table 7) in amounts comparable to those in leaves. Total nutrient concentration in berries was higher than that in tubers. Nitrogen application generally increased N concentration in the berries, but reduced berry K concentration significantly. The negative influence of K application on Mg and Ca are again well illustrated in Figures 6 and 7. These interactions strongly confirm K-Ca and K-Mg antagonism as reported by Terman and Allen (1974) and Marschner (1990). ACKNOWLEDGMENTS This work was done with the support of National Agricultural Research Organisation (NARO) and United States Agency for International Development (USAID)-Washington for a collaborative research on Solanum potato between Makerere University and Fort Valley State University, Georgia, USA. REFERENCES
Table 1. Soil characteristics at Namulonge Agricultural and Animal Production Research Institute and Kalengyere Research Station experimental sites
Soil textural name: Kalengyere = sandy loam; Namulonge = loam
Table 2. Effect of nitrogen and potassium application on dry matter and fresh tuber yield at Kalengyere Research Station during 1995 long rains (LR) and 1996 short rains (SR)
Values followed by the same letter or not followed by a letter
are not significantly different at P≤0.05. *=significant
at P≤0.05, **=significant at P≤0.01,
***=significant at P≤0.001. SHDM= Shoot dry matter
(%),TPP =number of tubers per plant, FTYield=Fresh tuber yield (t ha-1),
MTWT=Mean tuber weight (g), LDM=Leaf dry matter (%), TDM=Tuber dry matter (%)
Table 3. Effect of nitrogen and potassium application on dry matter and fresh tuber yield at Namulonge Research Institute during 1996 short rains and 1999 short rains (SR)
Values followed by the same letter or not followed by a letter
are not significantly different at P<0.05. *=significant at P<0.05, NS
= not significant at P = 0.05, BDM=Berry dry matter (%), TDM=Tuber dry matter
(%), FTYield=Fresh tuber yield (t ha-1), MTWT=Mean tuber weight (g),
TPP =number of tubers per plant
Table 4. Effect of nitrogen and potassium application on fresh tuber yield at Kalengyere Research Station and Namulonge Research Institute (NAARI) during 1999 short and long rains
Values followed by the same letter or not followed by a letter
are not significantly different at P<0.05. NS = not significant at P = 0.05,
TPP= number of tubers per plant, MTWT=Mean tuber weight (g), FTYield = Fresh
tuber yield (t ha-1)
Table 5. Leaf nutrient concentration in true potato seed mother plants as affected by N and K application at Namulonge Research Institute during 1996 short rains
Within treatments, in a column, values followed by the same
letter are not significantly different at P≤0.05.
Values that are not followed by a letter are not significant at P≤0.05.
NS = not significant at P = 0.05.
Table 6. Stem nutrient concentration in TPS mother plants as affected by N and K application at Namulonge during the short rains of 1996
Within treatments, in a column, values followed by the same
letter are not significantly different at P≤0.05.
Values that are not followed by a letter are not significant at P≤0.05
NS = not significant at P≤0.05
Table 7. Berry nutrient concentration in TPS mother plants as affected by N and K application at Namulonge during the 1996 short rains
Within treatments, in a column, values followed by the same letter are not significantly different at P≤0.05. Values that are not followed by a letter are not significant at P≤0.05, NA=not available; NS = not significant at P≤ 0.05 Table 8. Tuber nutrient concentration in true potato seed mother plants as affected by nitrogen and potassium application at Namulonge during 1996 short rains.
The following images related to this document are available:Line drawing images[cs01042k.gif] [cs01042h.gif] [cs01042g.gif] [cs01042b.gif] [cs01042i.gif] [cs01042m.gif] [cs01042f.gif] [cs01042d.gif] [cs01042l.gif] [cs01042e.gif] [cs01042j.gif] [cs01042a.gif] [cs01042c.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}