 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 9, No. 1, March 2001, pp. 175-184
M. Ebwongu, E. Adipala, S. Kyamanywa, C. K. Ssekabembe and A. S. Bhagsari1
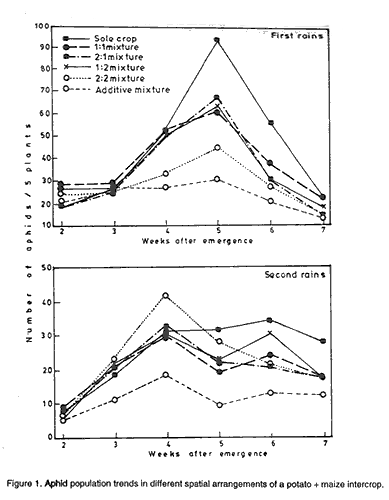
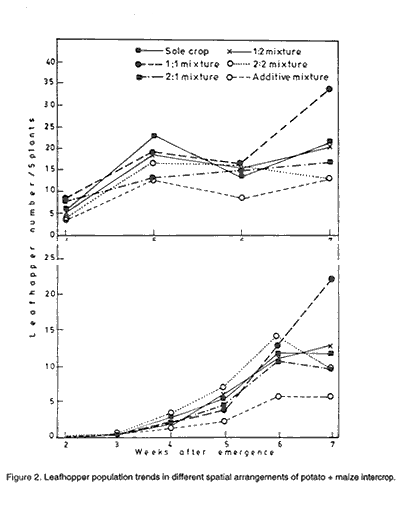
Code Number: CS01046 ABSTRACT A study to investigate the effect of intercropping maize (Zea mays L.) and potato (Solanum tuberosum L.) on the incidence of potato aphids and leafhoppers was conducted at Namulonge in Uganda during the two growing seasons of 1995. Three potato varieties and one maize variety were intercropped in six spatial arrangements; sole crops, 2:1, 2:2, 1:1, 1:2 potato: maize row arrangements and one additive mixture. Aphids and leafhopper infestations in the different spatial arrangements were similar at early stages but differed significantly later probably because increase in maize height with time and the associated effects had negative effects on pest establishment. Least aphid and leafhopper infestation were observed in the additive mixture. There was no significant influence of spatial arrangement on tuber damage although that tendency was for more damage in plots with high potato concentration. High relative humidity negatively affected aphids but not leafhoppers. Key Words: Additive mixtures, Myzus persicae, relative humidity, Solanum tuberosum, Zea mays RÉSUMÉ Une étude pour examiner les effets de la culture intercalaire du maiïs (Zea mays L.) et de la pomme de terre (Solanum tuberosum) sur l' incidence des aphides de la pomme de terre et des cicadelles a été conduite à Namulonge, Uganda pendant deux saisons culturales de 1995. Trois variétés de pomme de terre et une variété de maïs ont été associées dans des arrangements spatiaux de pomme de terre et maïs comme suit: monoculture, 2:1, 2:2, 1:1, 1:2 et un mélange additionel. Les infectations des aphides et des cicadelles étaient similaires dans les différents arrangements spatiaux aux premiers stades de croissance mais differaient significativement plus tards probablement à cause de l' augmentation avec le temps de la hauteur du maïs et les associations ont eu des effets négatifs sur l' établissement des pestes. Les plus faibles infestations des aphides et des cicadelles ont été observées dans le mélange additionnel. Les arrangements spatiaux n'ont pas eu d'effets importants sur les dégats des tubercules bien qu'une tendance était pour plus de dégats dans les parcelles avec une concentration élevée de pomme de terre. Une humidité relative élevée a affecté négativement les aphides mais pas les cicadelles. Mots Cles: Mélange additionnel, Myzus persicae, humidité relative, Solanum tuberosum, Zea mays INTRODUCTION In Uganda potato (Solanum tuberosum L.) is mainly grown in the highland areas of southwest Uganda and on the slopes of Mount Elgon in the east. These areas have rich volcanic soils with low temperatures and sufficient rainfall that are ideal climatic conditions for potato production. The production for 1998 was reported to be 190,158 tonnes on 28,269 hectares giving an average yield of 6.7 t ha-1. While reasonable potato yields may be obtained in the hilly areas of the tropics (Ahmad, 1977), which cover a small area in Uganda, high temperatures encountered in the lowland humid tropics place a major constraint on potato yields (Tizio, 1978; Menzel, 1983; Manrique, 1993). Approaches to improving potato production in the warm lowland tropics include the use of appropriate agronomic practices, correct choice of locally adapted varieties and the availability of cheap, high quality planting material (Midmore and Roca, 1992). Effect of intercropping on pest problems have been reviewed by many authors (Perrin, 1977; Taylor, 1977; Vandermeer, 1989; Ogenga-Latigo et al., 1993). A reduction of pests in intercrops have been reported in most reviews. Reduction of pests in intercropped potato has also been reported (Potts, 1990) and quantified (CIP, 1990; CIP, 1995). Other reports indicate variable results which depend on spatial arrangements of the intercrops. Intercropping has been used in some countries to modify the microclimate of potato to ameliorate or modify unsuitable climatic conditions including high temperatures and low soil moisture (Harverkort, 1989). This enables farmers to sandwich potato plants between slower growing cereals like maize. At low elevations mixed cropping is more prevalent with complex spatial arrangements of different crop species. In some areas potatoes are grown as a component of the slash - and - burn farming system, along with maize, beans and other crops (Horton, 1987). Like most other tuber crops, potato has many insect pests that attack the crop both in the field and in storage. The potato tuber moth, Phythorimaea operculella Zeller (Gelechiidae: Lepidoptera) is the most destructive field and storage pest of potato in warm climates (Kibata, 1982). The other pests include aphids, vectors of plant viruses including potato leaf roll virus (PLRV) and potato virus Y (Verma and Misra, 1975); leafhoppers (Raman and Radcliffe, 1992) and millepedes (Hill and Waller, 1988). Four genera of aphids attack potato, the most important being the green peach aphid, Myzus persicae Sulzer (Homoptera; Aphididea) (Kibata, 1982). This aphid is a worldwide pest of more than 30 plant families (Hollingsworth et al., 1990); and is responsible for transmission of over 60-90 viruses (Gibbs and Harrison, 1976; Behura, 1979), chief among them being potato leaf roll virus (PLRV) and potato virus Y (PVY). Losses in potato yield resulting from these viruses are estimated to be 20-50% and 40-85% for the respective viruses (Nagaich and Agrawal, 1969). They may also reduce yield and quality of tubers. When sufficiently abundant, aphids also inflict, on the growing plants, feeding damage that is reflected as reduced yield at harvest (Shands et al., 1972). The greatest damage from aphids occurs when high aphid populations cause reduced plant vigour and premature senescence (Hollingsworth et al., 1990). Certain species of leafhopper are known to serve as vectors for potato diseases. For example, in north America, aster leafhopper Macrosteles fascifrons transmits the aster yellows mycoplasma-like organism (AY-MLO) which causes purple top. Proper choice of intercropped species and appropriate agronomic practices reduce populations of various potato pests (Potts, 1988). Potato intercropped with wheat is less damaged by leaf miners because wheat attracts parasitoids of leaf miners (Raymundo and Alcazar, 1983). Reduction of pests in potato intercropped with maize, Oxalis tuberosa and other tuberous crops has also been reported in Peru (Raman and Radcliffe, 1992). However, not all intercrops reduce pests. For instance, Leroi et al. (1990) found that there was no difference in the number of eggs of Bruchidius atrolineatus deposited on sole cowpea and cowpea intercropped with millet. The objective of this study was to investigate the effect of intercropping potatoes and maize on two important pests (aphids and leaf hoppers) of potatoes in Uganda. MATERIALS AND METHODS The experiment was conducted during the first and second seasons of 1994 and 1995 at Namulonge (O° 32' N; 32° 35'E), at an altitude of 1150 m above sea level. Since the site is below 1500 m altitude it falls under mid-altitude elevation. Three potato genotypes of contrasting agronomic attributes and one maize variety (Longe 1) were used in the study. The characterists of these varieties are described in this volume (Ebwongu et al., 2001). The three potato varieties were grown in sole crops or potato + maize mixtures in the ratio 2:1, 2:2, 1:1, 1:2; the trial also had an additive mixture and sole maize. The treatments were in a split plot arrangement with the varieties in the main plots and spatial arrangement in the subplots. There were three replications each season. Weekly counts of aphids and leafhoppers were made on five potato plants per plot. These five plants were obtained from the inner sample rows in each plot. The number of aphids and leafhopper nymphs were counted. In addition to this, syrphid larvae and coccinellids were counted. Spider populations were estimated using pit-fall traps. Traps made from 0.5 litre oil container and having diameters of 7 cm were sunk into the soil with edges at ground level and filled with water and preservative to ¾ its depth. At weekly intervals the contents were emptied on a tea stainer to enable separation and identification of the insects. At harvest, 20 tubers of ware-size (50 + gm) were randomly picked, cleaned and inspected for insect tuber damage. The number of tubers damaged was then expressed as a percentage of 20. All the data obtained were subjected to Analysis of variance to separate significant treatment effects. Multiple regression was used to determine the relationship between various microclimate variables and yield or pest numbers. Square root (x + 0.5) transformation was applied to syrphid larvae numbers before analysis. RESULTS Aphids. Results of effect of different varieties and spatial arrangements on aphid infestation is illustrated in Table 1. Aphids numbers were significantly different (P<0.001) between the different varieties during both seasons. Kisoro consistently had significantly lower aphid numbers than Sangema and Victoria. The latter two varieties had almost equal infestation although their aphid population ranking changed between the seasons. Percentage plants infested by aphids followed a similar trend to that of aphid numbers per plant in the first season. In the second season the percentage aphid incidence on Victoria was more than for Sangema although severity was higher in the latter. Spatial arrangement significantly (P<0.05) influenced the infestation of potato by aphids (Table 1). The additive mixture consistently had the least infestation of 22.8 and 11.4 aphids/plot for the first and second seasons, respectively. This was followed by 2:2 potato/maize mixture in first season. In the second season the additive mixture was followed by 1:1 potato/maize mixture in having the lowest aphid incidence. Sole crop, on the other hand, was the most highly infested in both the first and second seasons (44.2 and 25.0 aphids/plot, respectively). In both seasons the effects of spatial arrangement x sampling time interactions were significant (P<0.05) on aphid infestation as shown in Figure 1. Populations were uniform in different spatial arrangements up to 2 (second season) and three (first season) weeks after planting (WAP). These periods were followed by significantly varied aphid populations among the spatial arrangements. Sole crop aphid population shot up to reach a maximum at 5 and 6 weeks after potato emergence in first and second seasons, respectively. In other spatial arrangements the populations build-up were generally low, and declined earlier than in sole crop. The additive mixture maintained the lowest aphid population during the entire sampling period. Aphid populations were influenced by climatic factors (Table 2). Aphid population had a significantly negative correlation with shade and relative humidity (R.H.) (r = -0.83, P = 0.004). However, shade was a more important factor affecting aphid population (r = -0.83, P = 0.008). Leafhoppers (Empoasca spp.). Infestation of varieties by the leafhoppers was not consistent during the study (Table 3). Leafhopper numbers on different varieties were only significantly different in the first season. However, Sangema consistently had higher number of leafhoppers per plot than both Victoria and Kisoro in both seasons. The effect of spatial arrangements on leaf hopper infestation was significant in both seasons with the 1:1 potato/maize mixture having the highest number of leafhoppers (Table 3) although they were not significantly higher than those in sole crop. The additive mixture, on the other hand, maintained significantly lower population of leafhoppers compared to the other treatments in both seasons. Leafhopper population was only significantly correlated with shade but not with relative humidity (R= -0.13, P = 0.11). Sampling time x spatial arrangement interactions for leafhopper infestation (Fig. 2) were significant in both seasons. Mixtures maintained relatively low leafhopper populations only early in the season. After 6 weeks after potato emergence leafhopper population in the 1:1 mixture shot up drastically. Natural enemy dynamics in potato/maize intercrop. The distribution of some natural enemies in the different spatial arrangement of potato and maize is shown in Table 4. Spiders were the most common insect predators on potato during the study. Predators responded in different ways to different cultivars and spatial arrangements. On average, 2.18, 2.28 and 1.88 spiders per trap were recorded on Kisoro, Sangema and Victoria, respectively in the first season. In the second season, 2.99, 3.74 and 3.72 spiders per trap were obtained from Kisoro, Sangema and Victoria, respectively. On the contrary, Syrphid larvae were attracted more to Victoria and least to Sangema in both seasons. Differences in numbers of Sryphid larvae on potato varieties were only significant in the second season. Spatial arrangements did not significantly affect the prevalence of the predators in both seasons. However, the 1:1, 1:2 and 2:2 mixtures had more predators than the sole crop (Table 4) and additive mixture consistently had the lowest number of syrphids. There were significant interactions between variety and spatial arrangements in number of syrphids in the second season. Tuber damage. Percent of insect-damaged tubers ranged from 1.6 to 7.1 and 2.4 to 10.9% in first and second seasons, respectively. Varieties did not significantly influence insect tuber damage (Table 5). Nevertheless in both the first and second seasons Kisoro had the least percent tuber damage, 2.67 and the 4.22%, respectively. The most affected variety in the first season was Sangema with 5.4% tuber damage. However, the rankings for Victoria and Sangema changed in the second season. Among the different spatial arrangements pure stands of potato showed the highest level of tuber damage in both seasons (Table 5). The other relatively high levels of attack were recorded in the 2:1 and 2:2 spatial arrangements. Least attack was recorded in the 1:1 mixture followed by 1:2 mixture. DISCUSSION This study has shown that pest incidence in different varieties and spatial arrangements varied significantly. Seasons also influenced pest numbers and their subsequent distribution in varieties and spatial arrangements. Pests build-up trends within seasons in different spatial arrangements appeared to have been affected by the growth of maize and its associated micro climatic changes. This is reflected in the significant cropping system x time interaction. There were varietal differences in respect to pest population, with consistently low foliar pest infestation and insect tuber damage on Kisoro. Leaves of Kisoro are pubescent and tiny, both of which are implicated in resistance to pests (Tingey and Laubengayer, 1981). Early maturing varieties (i.e., both Sangema and Victoria) had more aphids attracted to ageing leaves. Studies have shown that aged leaves contain soluble nitrates favourable for aphid growth (Kidd, 1986). The distribution of pests in different spatial arrangements might be attributed to a number of factors. The more apparent one is the micro-climate especially shade that increased as maize plants grew taller. Shading is known to change the quality of food (Kyamanywa and Tukahirwa, 1988) which aphids respond to by reducing colony sizes (Kidd, 1986; Ogenga-Latigo et al., 1993). This probably explains the proportionately bigger reduction in aphid numbers compared with the incidence of aphids in the first season when conditions were seemingly suitable for pest multiplication. Similar findings were reported by Rajat De and Singh (1979). In addition Kashyap and Bisnoi (1988) reported that high relative humidity, greater than 80-%, negatively affects pest population build-up. The other mechanisms explaining distribution of pests in the different spatial arrangements is the physical barrier effect of maize, which probably increased in importance as maize size and population increased in the mixtures. This phenomenon was clearly shown by the 2:2 and additive mixtures. Koestoni and Sastrosiswojo (cited by Potts, 1990) reported strip crop system as more effective than alternate rows in pest reduction which they attributed to barrier effects. However, high pest number later in the season in 1:1 and 1:2 mixtures is attributed to the trap effect of dense companion crop population that rendered pests especially leafhoppers unable to leave the plot. Leafhoppers are not significantly affected by microclimate so their populations continue growing once they have colonised the plants. So leafhopper populations were comparable to, or higher than those in sole crop resulted later in the season. Reduction in pest population could also have been due to slight increase in natural enemies in the intercrops (Risch, 1979). Some parasites are known to prefer dark environment available in the intercrops, while biodiversity in intercrops provide alternative food resources for the natural enemies. However, the similarity in numbers in sole and additive mixture suggests some response to prey populations too. This was apparent in the first season when more syrphid larvae were attracted to the sole crop than additive mixture. Natural enemy populations were insignificant and so played a minor role in influencing aphid populations. According to Risch et al. (1983) natural enemies play limited roles in reducing pest numbers in annual crop mixtures. In the current study, barrier effect was the most important mechanism in the mixtures. Cornelius and Barlow (1980) reported longer development stages of flower fly, Syrphus corollae when fed on Myzus persicae compared to Aphis pisum. This was attributed to the small size of M. persicae which provided low energy intake for the host and affected their reproduction. This observation may explain the small numbers of this predator that appeared in the study. The spiders seemed to be the most important predator available in potato/maize intercrop, as was also reported by Shands et al. (1972). This is probably because spiders are general predators and did not depend entirely on the few aphids that were present. Insect tuber damage was related to host concentration and ease of access to tubers. This is supported by the fact that more damage was realised in spatial arrangements with high potato populations. In addition, greater damage in sole crop in both seasons is attributed to wide soil cracks associated with better crop performance and high surface soil temperatures. ACKNOWLEDGEMENT The study was financed by the United States Agency for International Development (USAID) through a collaborative research grant to Makerere University and Fort Valley State College, Georgia, USA. REFERENCES
Table 1. Effect of spatial arrangement on aphid infestation of potato in a potato + maize intercrop during the first and second seasons of 1995
Table 2. Effect of potato + maize intercrop microclimate on pest populations
Table 3. Effect of spatial arrangement and potato variety on leafhopper infestation and incidence in a potato + maize intercrop during the first and second season of 1995
Table 4. Relative abudance of natural enemies in different spatial arrangements, 1995
Table 5. Effect of spatial arrangement and variety on potato tuber damage during the first and second season of 1995
Figure 1. Aphid population trends in different spatial arrangements of a potato + maize intercrop. Figure 2. Leafhopper population trends in different spatial arrangements of potato + maize intercrop. The following images related to this document are available:Line drawing images[cs01046b.gif] [cs01046a.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}