 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 9, No. 1, March 2001, pp.293-299
R. F. Karyeija, J, F, Kreuze1, R. W. Gibson3 and J. P. T. Valkonen2
Code Number: CS01060 ABSTRACT One sweetpotato breeding line resistant to sweetpotato feathery mottle virus (SPFMV) was infected by graft-inoculation with SPFMV and developed severe symptoms of sweetpotato virus disease, following co-inoculation with sweetpotato chlorotic stunt virus. Coat protein (CP) gene sequences from eight East African isolates of SPFMV and two West Africa isolates from Nigeria and Niger were determined. Phylogenetic analysis of sequences of these isolates and those of non-East African origin previously reported, showed that the East African SPFMV isolates form a separate cluster. The monoclonal antibody MAb 7H8 divided Uganda SPFMV isolates into serogroups, which differed in prevalence in different districts of Uganda and in two common sweetpotato cultivars. Key Words: Clustering, coat protein, East African SPFMV isolates, resistant sweetpotato cultivars, sweetpotato chlorotic stunt Crinivirus, West African SPFMV isolates Mab + isolates RÉSUMÉ Une lignée améliorée de patate douce résistante au virus à plume marbrée (feathery mottle virus: SPFMV) a été infectée par inoculation par greffage avec SPFMV et a développé des symptômes très sévères da la maladie virale de la patate douce, suivie d'une co-inoculation du virus de rabougrissement chlorotique de la patate douce. Les séquences des gènes d'une couche de protéines à partir de huit isolats East Africains de SPFMV et deux isolates West Africains du Nigeria et Niger ont été déterminés. Une analyse phylogénétique des séquences de ces isolates et ceux d'origine qui n'est pas de l'Afrique de l'Est prévieusement reportés, a montré que les isolates SPFMV de l'Afrique de l'Est forment un groupe séparé. Un anticorp monoclonale Mab 7H8 a divisé les isolats Ugandains SPFMV en deux séro-groupes, qui differaient en fréquences dans les differents districts de l'Uganda et dans cultivars communs de la patate douce. MotsClés: Groupement, couche de protéines, isolats SPFMV de l'Afrique de l'Est, cultivars résistant de patate douce, le virus de rabougrissememnt chlorotique de la patate douce, isolates Mab SPFMV de l'Afrque de l'Ouest INTRODUCTION Sweetpotato feathery mottle virus (SPFMV; family Potyviridae, genus Potyvirus) commonly infects sweetpotatoes (Ipomoea batatus) in Africa (Karyeija et al., 1998a) and elsewhere in the World (Moyer and Salazar 1989). It is transmitted by aphids in the non-persistent manner. In most East African sweetpotato cultivars, SPFMV causes no symptoms on the foliage (Gibson et al., 1997). However, dual infection with the unrelated whitefly-borne virus, sweetpotato chlorotic stunt virus (SPCSV; family Closterovirdae, genus Crinivirus) cause the severe symptoms known as sweetpotato virus disease (SPVD) (Schaefers and Terry, 1976). This is the main disease of the crop, characterised by small distorted leaves which are often narrow (strap-like) and crinkled with a chlorotic mosaic and /or vein-clearing and stunting of plants. SPVD-affected plants commonly produce less than half the tuberous root yield of symptomless ones. The variability of SPFMV in Uganda has not been studied and the availability of the monoclonal antibody Mab 7H8 (Hammond et al., 1992) allowed opportunity to study its serological variability. International Potato Centre (CIP) has also identified some sweetpotato clones including 420020 and 420026 known to be highly resistant and resistant, respectively, to infection by SPFMV in Peru (Fuentes and Salazar, 1996). These were used to examine the pathogenic variability of SPFMV isolates from SPVD-affected fields plants found in Uganda in SPFMV-resistant sweetpotato clones. Coat protein (CP) gene sequences of the Ugandan SPFMV isolates and those from elsewhere were also determined and reported elsewhere (Karyeija, 1999; Kreuze et al., 2000). MATERIALS AND METHODS Virus isolates. Sweetpotato plants with SPVD were obtained from farmers' fields in six districts in Uganda and transferred to a greenhouse at the University of Helsinki, Finland. Isolates of SPFMV were obtained from each plant using single aphids (Myzus persicae Sulz.) to inoculate I. setosa seedlings. The strain C of SPFMV originally isolated in North Carolina, U.S.A (Cali and Moyer, 1981), was obtained in desiccated leaves from CIP and was used to mechanically inoculate I. setosa. SPCSV was obtained by whitefly transmission from Ugandan SPVD-affected field plants (Gibson et al., 1998). Inoculation of resistant clones. Virus-free in vitro plantlets of sweetpotato clones 420020 and 420026 were obtained from CIP and grown in pots in a greenhouse at the University of Helsinki. In one experiment, batches of seven virus free plans of sweetpotato clones 420026 and 420020 were grafted with only SPFMV and three with a scion from the SPVD-affected sweetpotato clone from which SPFMV had been isolated. In a second experiment, two plants of the above-mentioned virus-free sweetpotato clones were grafted with a scion of I. setosa infected with only SPFMV as above, and two plants were graft-inoculated with a pair of scions, of which one was infected with SPFMV and the other with SPCSV. Inoculated sweetpotato plants were observed for symptoms and tested for SPFMV by NCM-ELISA 18-46 days after grafting. Any plant testing negative for SPFMV was grafted with scions obtained from virus-free I. setosa seedlings. Scions of I. setosa were observed for a further 28 days for possible symptom development and then tested for SPFMV by NCM-ELISA. Both experiments were done twice. Distribution of two SPFMV serotypes in Uganda. Leaves of sweetpotato
with SPVD symptoms collected from farmers' fields were tested by NCM-ELISA first
using a polyclonal antibody (PA b) to confirm the presence of SPFMV and then
using SPFMV Mab 7H8, to distinguish the monoclonal reactive serotype Mab + and
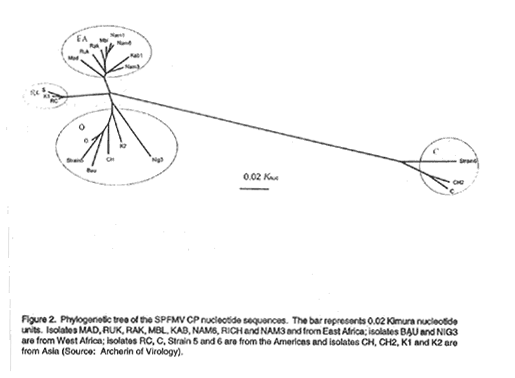
the non-reactive serotype Mab - of SPFMV. Coat protein sequencing and phylogenetic analysis. The CP encoding sequence was determined for seven Ugandan isolates (Kab1, Mb1, Nam 1, Nam 3, Nam 6, Rak and Ruk), and one SPFMV isolate each from Madagascar (Mad), Niger (Nig 3) and Nigeria (Bau) (Kreuze et al., 2000). Sequence alignments and phylogenetic analyses were done using the programs of the Wisconsin Package (Version 8, September 1994, Genetics Computer Group, Madison, WI) (Deveneux et al., 1984). The sequences obtained and those of other SPFMV CP sequences obtained from databases were aligned with the Pile Up Program (Gish and States 1993). The alignments were used to calculate phylogenetic distances with the Distances Program using the Kimura two-parameter model (Kimura, 1980). A phylogenetic tree was compiled with the neighbour-joining method (Saitou and Nei, 1987) using the Grow tree Program. RESULTS Serological analysis of SPFMV isolates. Isolates of SPFMV in Uganda fell into two groups: those which could and which could not be detected by the Mab 7H8 (Table 1). Detection of SPFMV serotypes in the field. The 108 SPVD-affected sweetpotato samples initially collected in Uganda in 1997, tested positive for SPFMV using Pab as also did a further 654 samples collected in four and five districts in 1997 and 1999 (Table 1). Mab + isolates were less common than the Mab - isolates in all districts except Mpigi in the 1997 sampling. The SPVD-resistant cultivar New Kawogo has become the predominant cultivar in parts of central Uganda and a separate survey was therefore done in fields of this and another less SPVD-resistant cultivar Tanzania, at Namulonge in September 1997 and April 1998. For both sampling dates, the plants of New Kawogo affected by SPVD also had similarly high proportions of Mab(+) SPFMV isolates. In contrast, only a few Mab(+) isolates were detected in the SPVD-affected plants of Tanzania (Table 2). Ability to overcome resistance. The SPFMV-resistant sweetpotato clones 420020 and 420026 were graft-inoculated with single isolates of Ugandan SPFMV and isolate C from the USA (Table 3). Neither the Ugandan isolates nor isolate C induced any symptom in plants of clone 420020, the leaves tested negative for SPFMV in NCM-ELISA and scions back-grafted to I. setosa seedlings did not induce symptoms. However, all Ugandan isolates induced a chlorotic mottle on leaves of the clone 420026 and these leaves tested positive for SPFMV in NCM-ELISA whereas isolate C failed to infect it. All plants of 420020 and 420026 graft-inoculated with both SPFMV and SPCSV became stunted, developed small distorted leaves with chlorotic mosaic symptoms characteristic of SPVD and tested positive for SPFMV by NCM-ELISA. Variability in SPFMV CP. The CP nucleotide (nt) sequences of eight SPFMV isolates from East Africa (Madagascar and five districts of Uganda) and two isolates from West Africa (Niger and Nigeria) were determined. Deduced CP amino acid (aa) sequences were compared to those of ten other SPFMV isolates from the database (Fig. 2). A total of 101 aa residues (32.1%) in the CP sequences showed variability, of which 46 were located in the N-terminal region. Overall the CP sequences were 75.6% identical at the nt level and 78.6% identical at the aa level (Kreuze et al., 2000). Phylogenetic analysis of the nt sequences split the SPFMV isolates into four groups (Fig. 2). The eight East African isolates from Uganda and Madagascar formed one group designated as Group EA, 95.8% and 96.5% identical at the nt and aa levels, respectively. Other groups were designated as:
DISCUSSION SPFMV is one of the most important Potyviruses, although it has been relatively poorly studied (Moyer and Salazar 1989; Karyeija et al., 1998a). Sweetpotato (I. batatas) originated from the Americas, and its current pan-tropical distribution is post-Columbus. However, the genus Ipomoea is naturally pan-tropical. Our findings that Ugandan SPFMV isolates form a single phylogenetic cluster by nucleotide sequence analysis and are unique in being able to infect CIP clone 420026 suggests either a unique evolutionary history of SPFMV in native Ipomoea spp in East Africa (Karyeija et al., 1998a; Karyeija et al., 1998b) and/or a single or limited introduction of SPFMV to East Africa. Despite East Africa isolates of SPFMV clustering by nucleotide sequences as a single group, Mab 7H8 identified two serotypes. Mab (-) isolates were predominant but Mab(+) isolates were found in all districts in both years. Taking the epitope recognised by Mab 7H8 as a marker of a "West African-type" CP, N-terminus (Hammond et al., 1992), the data suggested that cultivars traditionally grown in Uganda are less frequently infected with such isolates than the recently introduced resistant New Kawogo. Thus, the CP defining the two serotypes could be associated with host adaptation. That resistance to SPFMV in the sweetpotato clone 420020 (Fuentes and Slazar, 1996) was not overcome by Ugandan isolates, indicates that its resistance is broader than that of 420026. However, its resistance was overcome in the presence of SPCSV, plants developing SPVD. Consequently, selecting for resistance to SPFMV alone seems inappropriate, at least for Uganda where most damage by SPFMV is done in complex with SPCSV, and plants must be selected for resistance to both viruses. ACKNOWLDGEMENTS We thank H.J. Vetten, BBA Braunschweiz, for providing the SPVD-affected sweetpotato plants from West Africa. Grants from the Academy of Finland and FINNIDA, Finland (grant 34268), SAREC/SIDA, Sweden (grant SWE 1- 1997 - 141), and the British Department for International Development (DFID) are gratefully acknow-ledged. However, DFID can accept no responsibility for any information provided or views expressed. REFERENCES
TABLE 1. Prevalence of SPFMV serotypes in five districts of Uganda
TABLE 2. Detection of SPFMV isolation in the cultivars 'New Kawogo' and 'Tanzania' at Namulonge using the monoclonal antibody 7H8
aNumber of SPVD-affected plants that tested positively with
the Mab 7H8 in NCM-ELISA
TABLE 3. Graft inoculation of sweetpotato clones 420020 and 420026 with SPVD-affected sweet potato scions or scions of I. setosa infected with SPFMV from different locations in Uganda
Figure 1. The districts around Lake Victoria which form the main sweetpotato production area in Uganda. The sampling sites in this study were Rukungiri, Mbarara, Kabarole, Rakai, Masaka, Mpigi, Iganga and Mbale. The Kawanda Agricultural Research Institute (KARI) and the Namulonge Agricultural Research Institute are located in Mpigi District ca. 10 km and 20 km, respectively, North from Kampala. Figure 2. Phylogenetic tree of the SPFMV CP nucleotide sequences. The bar represents 0.02 Kimura nucleotide units. Isolates MAD, RUK, RAK, MBL, KAB, NAM6, RICH and NAM3 and from East Africa; isolates BAU and NIG3 are from West Africa; isolates RC, C, Strain 5 and 6 are from the Americas and isolates CH, CH2, K1 and K2 are from Asia (Source: Archerin of Virology). The following images related to this document are available:Line drawing images[cs01060a.gif] [cs01060b.gif] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}