
|
African Crop Science Journal
African Crop Science Society
ISSN: 1021-9730 EISSN: 2072-6589
Vol. 9, Num. 3, 2001, pp. 471-479
|
African Crop Science Journal, Vol. 9. No. 3, pp. 471-479
HETEROSIS AND COMBINING ABILITY IN A DIALLEL AMONG EIGHT
ELITE MAIZE POPULATIONS
M. NIGUSSIE and H. ZELLEKE 1
Nazret Research Centre, P. O. Box 436, Nazareth, Ethiopia
1Alemaya University of Agriculture, P.O. Box 138, Dire Dawa, Ethiopia
Received 3 April, 2000
Accepted 12 March, 2001
Code Number: cs01066
Abstract
Crossing maize (Zea mays L.) genotypes obtained from different sources
could result in better utilisation of hybrid vigor. Such heterotic response
is poorly exploited in the drought-stressed areas of Ethiopia, due to problems
in adaptation of some of introduced materials. This study was conducted to determine
the heterosis and combining ability of eight elite maize genotypes. The eight
parents were selected based on per se and top-cross performance. The
parents were crossed in diallel fashion. The resulting crosses and their parents
were evaluated in a randomised complete block design with three replicates at
three locations for two years in Ethiopia. The combined analysis of variance
showed that the mean square due to genotypes and general combining ability (GCA)
were significant (P=0.01) for all the six traits studied. However, the mean
square due to specific combining ability (SCA) was significant for days to tasselling,
days to silking, plant height, and grain yield. The magnitude of GCA was higher
than the SCA in all the cases indicating that additive gene action was more
important than non-additive in the inheritance of these traits. Mid-parent heterosis
occurred in varying degrees for the different traits. It was in the range of
- 11.6 to 21.9% for grain yield. DTP-2 C4 and Melkasa 92 DTP1 had significant
and positive GCA for days to tasselling, days to silking, plant height, and
grain yield. Hence, these parents can be used to develop intermediate maturing
varieties while AW-8047 significantly reduced (had negative GCA for) tasselling,
silking, and plant height without affecting grain yield indicating that AW-8047
can be used as source population to develop early varieties.
Key Words: Ethiopia, crosses, heterotic, genotypes, Zea mays
Résumé
Le croisement des génotypoes de maïs obtenus des differentes sources
pourraient aboutir à une meilleures utilisation de la vigeur des hybrides.
La réponse hétérotique est pauvrement exploitée
dans les régions à stress de séchéresse en Ethiopie,
suite aux problèmes d'adaptation de quelques materiels introduits.
Cette étude a été conduite pour déterminer l'hétérosis
et l'efficacité de combinaison de huit génotypes élites
de maïs. Les huit parents ont été séléctionnés
sur base des caractères intrénsiques et la performance supérieure
des croisés . Les parents ont été croisés selon
le modèle diallèle. Les croisés et leurs parents ont été
évalués dans un dispositif des blocs complètement randomisés
avec trois répétitions dans trois localités pendant deux
ans en Ethiopie. L'analyse combinée de la variance a montré
que le carré moyen des génotypes et l' efficacité
générale à la combinaison (GCA) étaient signifacatifs
(P<0.01) pour tous les six caractères étudiés. Cependant
le carré moyen de l'efficacité spécifique à
la combianaison (SCA) était significatif pour les jours au tasselling,
les jours au silking, la hauteur de la plante et au rendement grain. L'amplitude
de GCA était supérieure plus que SCA pour tous les cas suggérant
que l'action des gènes additifs était plus importante plus
que les non-additifs dans l'héritage de ces caractères.
L'effect hétérosis moyen du parent est apparu avec differents
degrés pour ces caractères. Il variait de - 11.6 à
21.9% pour le rendement grain. DTP-2 C4 et Melkasa 92 DTP1 avaient une GCA singificative
et positive pour les jours à tasselling, les jours a silking, la hauteur
de la plante et le rendement grain. Ainsi ces parents peuvent être utulisés
pour développer des variétés à maturité intermediaire
alors que AW-8047 a réduit significativemet (avait une GCA negative pour)
tasseling, silking , la hauteur de la plante sans affecter le rendement grain
indiquant que AW-8047 peut être utilisé comme source de population
pour développer des variétés précoces.
Mots Clés: Ethiopie, croisés, hétérotique,
génotypes, Zea mays
INTRODUCTION
In the drought-stressed areas of Ethiopia, maize (Zea mays L.) is one
of the most important food crops cultivated. It covers about 40% of the total
arable land in the country but contributes less than 20% to the total food
production (Mandefro et al., 1995). Low maize yields are mainly a result
of use of local poor yielding variety due to unavailability of suitable maize
varieties specifically high yielding hybrids. Fortunately, most farmers have
shown an increased interest in hybrid varieties that led Nazret Maize Programme
(NMP) to start work on developing hybrids for drought stressed environments
of the country.
Effective selection method for grain yield and other desirable traits requires
information on the magnitude of useful genetic variances present in the population,
in terms of combining ability and heterosis. A suitable means to achieve this
goal is the use of diallel mating systems, a method whereby the progeny performance
can be statistically separated into components related to general and specific
combining abilities (GCA and SCA). Combining ability analysis is one of the
powerful tools in identifying the better combiners which may be hybridised
to exploit heterosis and to select better crosses for direct use or further
breeding work.
Although such genetic studies have been made for maize grown
in other areas, little effort has been made to gather information on maize
improvement for drought stressed areas of Ethiopia. In 1995, a single isolated
study was done and thirty exotic maize populations were crossed to the open
pollinated cultivar Katumani and the resulting top crosses were evaluated
across locations. Results from 30 top crosses evaluation indicated that there
was high heterosis for most CIMMYT (International Maize and Wheat Improvement
Center) drought tolerant populations and negligible and/or negative heterosis
was recorded for African early maturing maize (Mandefro et al., 1997).
Further studies are needed. The objective of this study was to determine heterosis
and estimate combining ability of eight elite maize populations.
MATERIALS AND METHODS
Based on per se and top-cross performance, eight elite maize populations
were chosen to form the diallel at Melkasa (Ethiopia) during the 1997 off-season
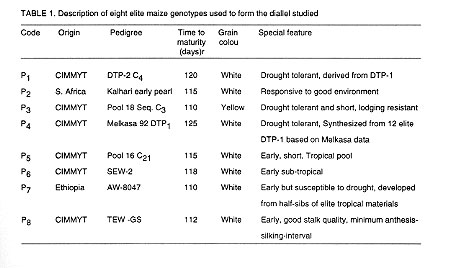
(December-April). Detailed descriptions of the genotypes are presented in
Table 1. Fresh seeds of the eight parents
were planted in 10 rows of 5 m length for each cross combination. All possible
28 crosses were made in both directions using bulk pollen. Seeds of each cross
and reciprocal (60 cobs for each cross) were bulked for use in trials. The
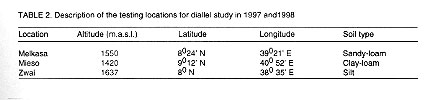
resulting 28 crosses and the eight parents were evaluated at Melkasa, Mieso,
and Zwai (locations are described in Table
2) in 1997 and 1998 main seasons (June-November). The experimental design
was randomised complete block with three replicates. The plot consisted of
two 5 m rows spaced at 75 cm apart. Two plants per hill were planted first
and then thinned to one plant per hill with 25 cm between hills, giving a
final plant density of 53,333 plants per hectare. Phosphorus (P2O5)
at a rate of 46 kg ha-1 was applied at the time of planting while
N at a rate 41 kg ha-1 was applied of which 18 kg ha-1
was applied at planting, and the remaining 23 kg ha-1 36 days after
emergence. Planting was done at the onset of the rainfall (in June). Plots
were weeded on the 25th and 40th days after emergence.
Plots were harvested and shelled manually. The shelled grain yield (kg ha-1)
was calculated after adjusting the moisture content to 12.5 percent.
Data were collected for days to tasselling, days to silking,
plant height, number of ears per plant, thousand seed weight and grain yield.
Analysis of variance was computed first for each location separately (data
not shown) and then combined across locations. Each season at a given site
was considered as a different location (i.e., the three locations by two years
were considered as six locations). Analysis of variance and diallel analysis
was done using a computer software suitable for Griffing's Model I,
method 2, which involves parents and one set of crosses (Griffing, 1956).
The mathematical model for the combining ability is
X ijkl = μ + gi + gj + Sij
+ bk + eijkl
where X ijkl is the performance of the cross between ith
and jth genotypes in the kth replicate at lth
location, μ is the population mean, gi(gj) is the GCA effect, Sij
is the SCA effect, such that Sij = Sji, eijkl
is the effect peculiar to the ijklth observation. The restrictions
ági = 0, and ásji
+ sii = 0 (for each i) were imposed. The percentage of mid-parent
and better-parent heterosis was computed as outlined in Falconer (1981)
RESULTS AND DISCUSSION
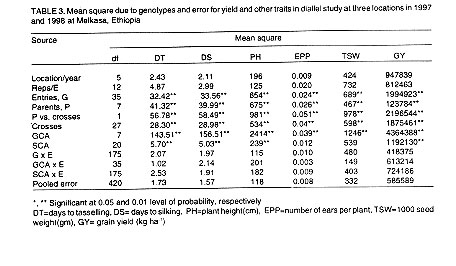
Analysis of variance and genotypic mean. The combined analysis of variance
showed that the mean square due to genotypes and general combining ability
(GCA) were highly significant (P=0.01) for days to tasselling, days to silking,
plant height, number of ears per plant, 1000 seed weight and grain yield (Table
3). The mean square due to specific combining ability (SCA) was significant
for days to tasselling, days to silking, plant height and grain yield (Table
3).
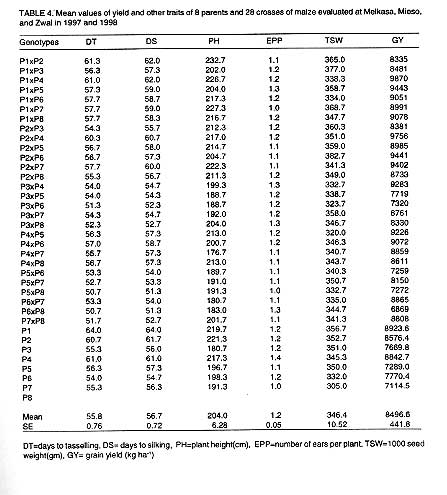
The mean performance of parents and their hybrids across locations are presented
in Table 4. The highest grain yield was
9870 kg ha-1 in P1 x P4 (DTP-2 C4 x Melkasa 92 DTP-1)
followed by P2 x P4 (Kalhari Early P. x Melkasa 92 DTP1) and P1 x P5 (DTP-2
C4 x Pool 16 C21) with grain yield of 9756 and 9443
kg ha-1, respectively. The lowest grain yield (6869 kg ha-1)
was recorded for P6 x P8 (SEW-2 x TEW-GS). Among the parents, the best yield
was obtained from P1 (DTP-2 C4) while the least was from P7 (AW-8047).
Thousand seed weight ranged from 383 gm for P2 x P6 (Karlhari early P. x SEW-2)
to 324 gm in P4 x P5 (Melkasa 92, DTP1x Pool 16 C21
x SEW-2) for the crosses, and it varied from 357 for P1 (DTP1 -C4)
to 305 g obtained from P7 (AW-80 47) for the parents. The number of ears per
plant ranged from 1.4 for P4 (Melkasa 92 DTP1), to 1.0 for P1 x
P7. Number of ears per plant, one of the criteria used to measure stress tolerance
was greater than or equal to 1.0 for all genotypes may be due to the uniform
and reliable rains during the execution of the experiment.
Plant height was in the range of 233 (P1 x P2) and 177 cm (P4 x P7) while
the highest ear height was 117 cm (P1 x P7) and the lowest was 73 cm obtained
from P5 x P6 (Pool 16 C21 x TEW-GS). The maximum number of days
to silking was 62.0 days obtained from P1 x P2 or P1 x P4 and the minimum
was 51.3 days obtained from P6 x P8 or P5 x P8 for the crosses while it was
in the range of 64.0 (P1) to 51.0 days (P8) for the parents. Number of days
to tasselling followed a similar trend as that of silking.
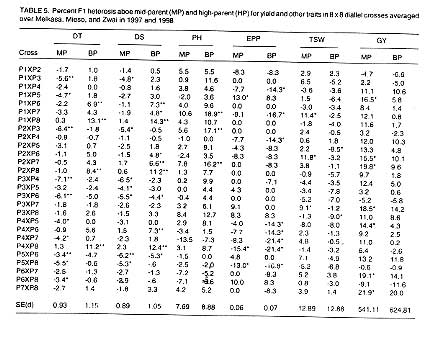
Heterosis. The values of heterosis over the mid-parent (MP) and the
high-parent (HP) are presented in Table 5.
Different hybrids showed significant heterosis for the traits studied. Eleven
crosses expressed negative and significant mid-parent heterosis for days to
tasselling, whereas heterosis above the high- parents was obtained for four
crosses for the same trait. Heterosis above the mid- and high-parent for days
to tasselling were reported by Vasal et al. (1992a). Eight crosses
had significant and positive heterotic values of better-parent for days to
silking. Kebede Malatu (1989) and Vasal et al. (1992b) reported better-parent
heterosis for days to silking. The percentage heterosis over the better-parent
ranged from 0.0% to 18.9% (P1xP7) for plant height. Significant and negative
mid-parent heterosis was observed in two crosses for number of ears per plant.
Crosses that exhibited negative heterosis had gene combination that reduce
the number of ears per plant. Three crosses showed positive and significant
heterosis above the mid-parent for 1000 seed weight while other two crosses
showed negative and significant heterosis for the trait. This is in agreement
with earlier findings of Shewangizaw (1983) who found heterosis above the
better-parent only in few crosses for 100 seed weight. Seven crosses showed
significant and positive heterosis above the mid-parent for grain yield. Heterosis
above the high-parent for grain yield was recorded only for P7 x P8. The expression
of grain yield above the mid-parent and high-parent has been reported by several
investigators (Beck et al., 1990; Vasal et al., 1992a).
In general, heterosis or hybrid vigour was more frequent in days to tasselling
and days to silking than for other traits studied in this work. Heterosis
above high-parent was more variable than the mid-parent values ranging from
-4.4% (P3 x P6) to 14.3% (P1 x P8) for days to silking indicating that earliness
was highly enhanced in P3 x P6 while lateness was favoured in P1 x P8, and
the choice from the two hybrids depends on the environmental situation for
which the hybrid is targeted.
Crosses, which had divergent parents with respect to growth habit, exhibited
maximum heterosis relative to the better-parent. P7 x P8 that showed maximum
heterosis (21.9%) for grain yield, had parents that were different in their
growth habit. This observation confirms that heterosis in maize tends to increase
with increase in divergence among parents with respect to geographical origin,
seed size and growth characteristics. Gallais (1984) made observations that
collaborate with this principle and he also stated that extreme diversity
or very related parents decreased heterosis.
Combining ability. Mean squares due to general combining ability (GCA)
were significant for all traits measured (Table
3), whereas the mean squares due to specific combining ability (SCA) were
significant for days to tasselling, days to silking, plant height, and grain
yield implying that these traits are governed by both additive and non additive
gene action. For plant height, both additive and non-additive gene actions
were equally important in this study. Similar results were reported by Zambezi
et al. (1986).
Several studies involving the inheritance of various quantitative traits
in maize have revealed the predominance of additive gene action (Shewangizaw,
1983; Stangland et al., 1983; Zambezi et al., 1986). This has
breeding implications, since additive genetic variation can effectively be
exploited by simple recurrent selection. The performance of parents could
be used to predict the performance of crosses. Thus, parents with good general
combining ability (GCA) and per se performance can be crossed to develop
high yielding composites that can be used directly for recommendation or for
further breeding work.
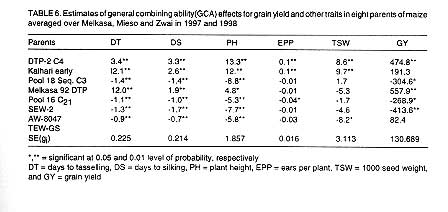
Estimates of GCA effects of parents for various traits are presented in
Table 6. Kalhari early pearl, DTP-2 C4
and Melkasa 92 DTP1 showed positive and significant values of GCA
for days to tasselling, silking and plant height. The highest GCA value for
all the three traits was obtained from DTP2 C4. All
the remaining parents exhibited negative and significant GCA effects for days
to tasselling, and silking. Three parents had negative and significant GCA
values for plant height for example, AW-8047. DTP-2 C4, and Kalhari
early pearl, were good combiners for number of ears per plant while Pool 16
C21 was poor combiner for number of ears per plant.
Good combiners for 1000 seed weight were DTP2-C4 and
Kalhari early pearl. Variety AW-8047 showed significant and negative GCA value
for 1000 seed weight. Positive and significant GCA effects were exhibited
by DTP-2 C4 and Melkasa 92 DTP1 for grain yield. These two parents
were also good combiners for most of the traits studied. Negative and significant
GCA effect were manifested by Pool 18 seq C3, Pool 16 C21,
SEW-2 and TEW-GS for grain yield.
In this study, DTP-2 C4 was a good combiner for all traits. On the other
hand, Pool 18 seq C3, Pool 16 C21, SEW-2, AW-8047, and
TEW -GS significantly decreased days to tasselling and silking. The latter
parents also decreased plant height significantly. Pool 18 seq C3
significantly reduced grain yield. Pool 16 C21 reduced significantly
the number of ears per plant and grain yield while AW-8047 significantly reduced
1000 seed weight. The present results agree with the work of Hallauer and
Miranda (1988).
In general, the parent DTP-2 C4, Kalhari early pearl and Melkasa
92 DTP1, contributed to lateness in tasselling and silking, and
to increasing plant height and grain yield (except Kalhari early pearl). It
is common for maturity and plant height to be associated with grain yield
(Hallauer and Miranda, 1988). Pool 18 seq C3, SEW-2, AW-8047, Pool 16 C21
and TEW-GS contributed earliness in days to tasselling and silking. Pool 18
seq C3, Pool 16 C21, SEW-2 and TEW-GS reduced grain
yields significantly for the reason that earliness has inverse relation with
grain yield (Bolanos and Edmeades, 1993). However, AW-8047 significantly reduced
days to flowering and maturity but did not affect grain yields. This parent
may have a specific trait that stimulates the physiological activities in
the plant system that contribute to higher grain yields (high efficiency in
conversion rate of assimilate per unit time).
Parents that are relatively late in maturity and possess drought tolerance
characteristics can be used to develop varieties/hybrids that are intermediate
in maturity and useful for April planting in Ethiopia. Early maturing parents
are meant to develop early varieties for June planting in the drought stressed
areas of the country (where the growing season is so short).
P4 x P8 was the only cross that had significant and positive SCA (data not
shown) effect for days to tasselling, implying that lateness in tasselling
was enhanced in this particular cross. On the contrary, P1 x P3, P2 x P3,
P3 x P4, and P5 x P8 exhibited negative and significant SCA values, indicating
that these crosses tasselled earlier than what would have been predicted based
on their parental performance. Crosses P2 x P7, P4 x P6 and P4 x P8 had significant
and positive SCA effects for days to silking showing that parents involved
in these crosses enhanced lateness in female flowering. Other crosses, such
as P1 x P3, P3 x P4, P3 x P6, P5 x P7 and P5 x P8, exhibited negative and
significant SCA indicating that these crosses had shorter number of days in
female flowering than the expected mean performance of their parents. In general,
P3 and P5 enhanced earliness in male and female flowering when crossed to
any other parents used in this study and may be used as good sources for earliness.
Crosses P1 x P7 and P2 x P7 exhibited significant positive SCA effects for
plant height, while P4 x P7 showed significant and negative SCA effect.
The two crosses that showed significant and positive SCA values for grain
yield were P1 x P5 and P6 x P7, indicating that these two crosses combined
well to give higher grain yields than the mean performance of their respective
parents. Manifestation of good SCA may not be expected, since the parents
involved in producing the crosses were broad-based populations. Several authors
reported that good GCA effect is obtained from broad based parents (pools,
populations, half sibs, fulsibs, composites), while good SCA is exhibited
by narrow based (inbred lines) parents upon crossing.
CONCLUSIONS
The results of the study have demonstrated the importance of diallel analysis
in detecting heterosis and identifying parents with general and specific combining
abilities that help develop hybrids suitable for desirable traits. Additive
gene action predominated in all characters studied. The dominance of additive
gene action in the inheritance of quantitative traits can be effectively exploited
by simple recurrent selection. Kalhari early pearl, DTP-2 C4 and
Melkasa 92 DTP1 showed positive and significant values of GCA for days to
tasselling, and silking. These parents can therefore be used to develop intermediate
maturing varieties to be planted in areas receiving bi-modal rainfall pattern
(for April planting). Variety AW-8047 significantly reduced days to tasselling,
silking and plant height without affecting the grain yields and hence AW-8047can
be used in developing early maturing varieties.
ACKNOWLEDGEMENT
We are deeply indebted to National Maize Programme staff who assisted in the
evaluation of these materials. We also thank African Crop Science Journal Reviewers
for their comments and corrections.
REFERENCES
-
Beck, D.L., Vasal,S.K., Crossa, J. 1990. Heterosis and combining ability
of CIMMYT's tropical early and intermediate maturity maize germplasm.
Maydica 35:279-285.
-
Bolanos, J. and Edmeades, G.O. 1993. Eight cycles of selection for drought
tolerance in low land maize II. Response in reproductive behaviour. Field
Crops Research 31:253-268.
-
Falconer, D.S. 1981. Introduction to quantitative genetics. Longman,
New York.
-
Gallais, A. 1984. An analysis of heterosis Vs inbreeding effects with
an autotetroploid cross fertilized plant Medicogo Sativa L. Genetics
106:123-137.
-
Griffing, B. 1956. Concept of general and specific combining ability
in relation to diallel crossing systems. Australian Journal of Biological
Science 9:463-493.
-
Hallauer, A.R. and Miranda, J.B. 1988. Quantitative genetics in maize
breeding. 2nd (Ed.). Iowa State University Press, Ames. U.S.A.
-
Kebede Mulatu. 1989. Manifestation of heterosis and the nature of inheritance
of traits among reciprocally crossed inbred lines and population of maize
(Zea mays L.). M.Sc. Thesis, Alemaya University of Agriculture. Ethiopia.
-
Mandefro Nigussie, Gezahegne Bogale and Benti Tolessa. 1997. Top cross
performance and heterotic patterns of 31 maize populations in the drought
stressed areas of Ethiopia. In: Proceedings of the Crop Science Society
of Ethiopia Sebil 8:26-27.
-
Mandefro Nigussie, Gezahegne Bogale and Gelana Sebokesa.1995. Maize breeding
for
drought stressed areas of Ethiopia. In: Proceeding of the 25th Anniversary
of Nazret Agricultural Research Center: 25 years of experience in low land
crops research, 20-23 September 1995. Habtu Asefa (Ed.), pp. 68-74.
Nazareth, Ethiopia.
-
Shewangizaw Abebe. 1983. Heterosis and combining ability in 7x7 diallel
cross of selected inbred lines of maize (Zea mays L.). M.Sc. Thesis,
Addis Ababa University, Ethiopia.
-
Stangland, G. T., Russell, W.A. and Smith, O.A. 1983. Evaluation of the
performance and combining ability of selected lines derived from improved
maize populations. Crop Science 18:224-226.
-
Vasal, S.K., Srinivasan, F., Gonzalez, C., Han, G.C. and Shivaji Pandey
1992a. Heterosis and combining ability of CIMMYT's Tropical x subtropical
maize germplasm. Crop Science 32:1483-1489.
-
Vasal. S.K., Srinivasan, G., Beck, D.L., Crossa, J, Pandey,S., De Leon,
C.1992b. Heterosis and combining ability of CIMMYT's tropical late
white maize germplasm. Maydica 37:217-223.
-
Zambezi, B.T., Harner, E.S. and Martin, F.G. 1986. Inbred lines as testers
for general combining ability in maize. Crop Science 26:908-910.
© Copyright 2001, African Crop Science Society
The following images related to this document are available:
Photo images
[cs01066t3.jpg]
[cs01066t5.jpg]
[cs01066t6.jpg]
[cs01066t4.jpg]
[cs01066t2.jpg]
[cs01066t1.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}