
|
African Crop Science Journal
African Crop Science Society
ISSN: 1021-9730 EISSN: 2072-6589
Vol. 9, Num. 3, 2001, pp. 527-535
|
African Crop Science Journal, Vol. 9. No. 3, pp. 527-535
STUDIES ON THE INTERACTION BETWEEN RALSTONIA SOLANACEARUM
(SMITH) AND MELOIDOGYNE SPP. IN POTATO
E.M. ATEKA, A.W. MWANG'OMBE and J.W. KIMENJU
Department of Crop Protection, University of Nairobi, P.O Box 30197, Nairobi,
Kenya
Received 9 March, 1999
Accepted; 30 March, 2001
Code Number: cs01072
ABSTRACT
A survey was undertaken to determine population density of Meloidogyne
juveniles (J2) in 90 fields randomly selected from three potato-producing
districts in Kenya namely Nyeri, Meru and Nyandarua. Mean nematode densities
were highest (45) in samples collected from Nyeri and Nyandarua and lowest
(31) in soil samples collected from Meru. The reaction of 15 potato cultivars
to Meloidogyne incognita was determined under greenhouse conditions.
Plants were inoculated with 6000 eggs and second-stage juveniles each. Highly
significant (P=0.01) differences were observed between the cultivars. Galling
index was highest (5.5) in cv. KP93739.26 and lowest (1.9) in cv. Nyayo. All
cultivars supported nematode reproduction with the highest (5.0) egg mass
index being recorded in cultivars KP93739.26, Kerr's Pink, Desiree CIP-800048,
KP92633.26 and B53. No cultivar exhibited immunity. The relationship between
infection by root-knot nematodes and severity of bacterial wilt in three potato
cultivars with varying levels of resistance to bacterial wilt namely Asante
CIP 381381.20 (susceptible), B53 (moderately susceptible), and Kenya Dhamana
(resistant), was investigated in a greenhouse experiment. Bacterial wilt was
more severe in plants infected with both pathogens as compared to plants infected
with R. solanacearum alone.
Key Words: Egg mass index, galling index, incidence, Meloidogyne,
severity
Résumé
Une enqueté a été enterprise pour déterminer la
densité de la population de jeunes Meloidogines (JM) dans 90 champs
aléatoirement séléctionnés dans trois districts
producteurs de la pomme de terre au Kenya à savoir Nyeri, Meru et Nyandarua.
Les moyennes des densités des nématodes étaient plus élevées
(45) dans les échantillons collectés dans Nyeri et Nyandarua et
plus faibles (31) dans les échantillons de sols collectés dans
Meru. La réaction de 15 cultivars de pomme de terre au Meloidogine
incognita a été déterminée dans les conditions
de serre. Les plantes étaient inocullées avec 6000 oeufs et le
deuxième stade juvénile chacun. Des differences hautement significatives
(P=0.01) ont été observées entre les cultivars. L'indexe
de galles était plus élevé (5.5) chez le cultivar KP93739.26
et plus faible chez le cultivar Nyayo. Tous les cultivars ont supporté
la production de nématodes avec l'indexe de masse des oeufs plus
élevé chez les cultivars KP93739.26, Kerr's Pink, Desiree
CIP-800048, KP92633.26 et B53. Aucun cultivar n'a montré d'
immunité. La relation entre l'infection par les nématodes
des noeuds des racines et la séverité du mildiou dans trois cultivars
de pomme de terre avec different niveaux de résistance au mildiou à
savoir CIP 381381.20 ( sensible), B53 ( modérement sensible), et Kenya
Dhamana (résistant), a été étudiée dans un
essai en serre. Le mildiou était plus sévère dans les plantes
infectées par les deux pathogènes par rapport aux plantes seulement
infectées par R. solanacearum.
Mots Clés: Indexe de masse des oeufs, indexe de galles, incidence,
Meloidogyne, sévérité
INTRODUCTION
Potato (Solanum tuberosum L.) is the fourth most important food crop
in the world after wheat, rice and maize with its production roughly representing
half the world's annual output of all root crops and tubers (FAO, 1995).
However, its cultivation is limited largely by diseases and pests (Hayward
and Hartman, 1994).
Bacterial wilt was first reported in Kenya in the1940s and, since then, it
has spread to most potato growing areas (Michieka, 1993). Over 50% yield losses
have been reported in the country with occasional losses of up to 7 % on seed
potato (Ajanga, 1993). The disease is a major constraint to potato production
particularly because its management is difficult (Sequeira, 1992).
Root-knot nematodes (Meloidogyne spp.) are another major limiting factor
to potato production worldwide (Bird, 1981). Their feeding reduces growth
vigour and causes blemishes on tubers thus reducing their marketability (Sijmons
et al., 1994). Losses resulting from root-knot nematode damage alone
can reach 25% depending on the cultivar and prevailing environmental conditions
(Mai et al., 1981).
Meloidogyne spp. occurs commonly in warm regions and, are therefore
not a worldwide problem on potato because potatoes are mainly grown in cool
regions (Evans and Trudgill, 1992). However, with increasing land pressure,
production is expanding to the warmer lowlands where root-knot nematodes are
undoubtedly troublesome.
The use of resistant cultivars has several advantages over other methods
of reducing nematode populations; it requires little or no technology, it
is cost effective and does not leave toxic residues (Trudgill, 1991). Nematicides,
besides leaving toxic residues in the environment, provide partial control
(Brown et al., 1989).
The influence of root-knot nematodes on the development and severity of
bacterial wilt has been elucidated (Akiew et al., 1991). Plants infected
with both Meloidogyne spp. and Ralstonia solanacearum have been
found to express wilt symptoms earlier with greater severity than those infected
with R. solanacearum alone (Johnson and Powell, 1969; Akiew et al.,
1991). Sitaramaiah and Sinha (1985) have suggested that the nematode induced
stress is more important as a wilt-triggering factor than as a wounding agent.
Whereas the reaction of most commercial potato cultivars to major diseases
such as late blight (causal agent Phytophthora infestans), and bacterial
wilt, Ralstonia (Pseudomonas) solanacearum, has been established, their
reaction to root-knot nematodes is largely unknown. Additionally, potato reported
as resistant in a given location may not be resistant to the same root-knot
nematode species when grown in another location. In Kenya, the distribution
and density of root-knot nematodes in potato growing areas is also unknown.
Even though infection by Meloidogyne spp. is known to predispose
potato plants to R. solanacearum, information on occurrence of the
two pathogens and their interaction in Kenya is meagre. In addition, information
is lacking on the impact of Meloidogyne infection on wilt expression
in potato cultivars with varying levels of resistance to R. solanacearum.
The objectives of this study therefore were to (a) determine the occurrence
of root-knot nematodes in Kenya, (b) assesse the interaction between bacterial
wilt pathogen R. solanacearum and root-knot nematode Melodogyne
spp. (c) determine the reaction of potato cultivars to M. incognita.
MATERIALS AND METHODS
Population densities of Meloidogyne spp. in potato fields. Thirty
potato fields in Nyeri, Nyandarua and Meru district were randomly selected
at intervals of 3-5 kilometers along the roads. The sampling procedure for
nematode assessments was adopted from Dropkin (1980). Each potato field was
divided into four blocks. From each block, five soil samples were taken from
the rhizosphere of potato plants along a zigzag path. The top 5-cm soil layer
was scrapped off before a sample of soil was obtained from a depth of 5-20
cm. Twenty soil samples were taken from each farm and mixed thoroughly before
3 kg of the composite sample was taken. Nematode extraction was done by the
sieving and filtering technique (Hooper, 1990). Two-200 cm3 soil
sub-samples were taken from the composite sample and placed in a bucket to
which about 5 litres of water was added. The resulting suspension was stirred
and passed through a 710 μm-aperture sieve nested on top of a
45 μm-aperture sieve. The residues on the 710 μm-aperture
sieve were discarded while those on the 45μm-aperture sieves
were backwashed into a beaker and then poured onto double milk filters supported
on a screen standing in a shallow dish. Water was added into the dish until
it was just touching the milk filters. Nematodes were allowed three days to
move from the suspension, through the milk filters, into the water beneath.
The resulting nematode suspension was concentrated by draining off excess
water through four 45 μm-aperture sieves. The nematode suspension
was adjusted to a volume of 10 ml by adding or siphoning excess water after
allowing the nematodes to settle at the bottom of the vials. One-ml aliquot
of a well-mixed suspension was drawn and pippeted into a counting slide and
counting done under a light microscope. Counting was repeated in three aliquots
to improve accuracy.
Resistance of potato to Meloidogyyne incognita.
Fifteen potato cultivars were evaluated for resistance to M. incognita
in the greenhouse. The cultivars were Nyayo, Desiree CIP-800048, Roslin Tana,
Kerrs Pink LB-5, Golof (Dutch Robjin), B53, Tigoni CIP-381381.13, Rutuku CIP-720097,
Asante CIP-381381.20, Kenya Dhamana CIP- 800228, Mauritius Clone, KP93739.26,
KP92633.26, Furaha and Cruza148 CIP-720118. Tomato (Lycopersicon esculentum
L.) cv. Cal J was included as a susceptible control since there was no potato
cultivar with known reaction to M. incognita. Potato tubers were sown
in 20-cm diameter plastic pots containing heat-sterilized potting mixture
of soil, sand and ballast at the ratio of 3:2:1. Inoculum consisted of 6000
eggs and second-stage juveniles of M. incognita suspended in 10 ml
sterile distilled water. The inoculum was pipetted into 4-cm deep indentations
made in the soil medium at the root zone of each plant, one-week after planting.
Treatments were arranged in a completely randomised design with eight replicates.
Control plants were treated with water. Plants were watered as often as necessary
and fertilised with 50 g of diamonium phosphate.
The experiment was terminated eight weeks after soil infestation. Plants
were gently uprooted from pots and roots washed free of soil. Determination
of root damage was based on a scale of 0-10 where: - 0 = complete and healthy
root system, no infestation, 1 = very small galls only detected upon close
examination, 2 = small galls as in 1, but more numerous and easy to detect,
3 = numerous small galls, function of roots seriously affected, 4 = numerous
small galls, some big galls, majority of galls still functioning, 5 = 25%
of the root system severely galled and not functioning, 6 = 50% of root system
severely galled and not functioning, 7 = 75% of root system severely galled
and lost for production, 8 = no healthy roots, plant still green , 9 = the
completely galled root system is rotting, plant was dying, 10 = plants and
roots dead (Zech, 1971).
Egg mass index (EMI) was used to determine the ability of each cultivar
to support nematode reproduction. Galled roots were washed free of soil and
then dipped in 1% solution of Phloxine B to stain the egg masses. An egg mass
index scale of 0-5, where 0= 0, 1 = 1-2, 2 = 3-10, 3 = 11-30, 4 = 31-100,
and 5 = >100 egg masses per root system, was used (Taylor and Sasser, 1978).
Effect of Meloidogyne incognita on the expression of bacterial wilt
of potato. A greenhouse test to determine the effect of Meloidogyne
incognita infection on the expression of bacterial wilt in potato cultivars
with varying levels of resistance to bacterial wilt was conducted from August
to October 1998. Three potato cultivars namely Asante CIP-381381.20 (very
susceptible), B53 (moderately susceptible), and Kenya Dhamana CIP- 800228
(resistant) were used in this test. Certified tubers of these cultivars were
sown in pots containing heat-sterilised soil: sand (2:1) mixture. Treatments
included plants inoculated with (i) R. solanacearum without root wounding
(severing) (ii) R. solanacearum with root wounding by the procedure
described by Winstead and Kelman (1952) (iii) Meloidogyne incognita
and R. solanacearum without root wounding (iv) M. incognita
and R. solanacearum with root wounding. Controls included (a) plants,
which were not inoculated with any pathogen but with wounded roots (b) plants
which were not inoculated with any pathogen but with root wounding.
Nematode inoculum was extracted from galled tomato roots following the procedure
described by Omwega et al. (1988). Galled tomato roots were obtained
from two-month old tomato plants that were inoculated with M. incognita
when they were two weeks old. The roots were washed free of soil and thereafter
dipped in distilled water for about 1 hr to allow any other form of nematodes
to move out into the water leaving behind M. incognita eggs. The roots
were then dipped in sterile distilled water that was being aerated by an aquarium
pump. The eggs hatched into juveniles, which moved into the water in about
5-10 days. Inoculum consisted of 6000 Meloidogyne J2 suspended
in 10 ml sterile distilled water. Inoculation was done by pippetting the inoculum
into the soil in indentations made in the root zone of each plant two weeks
after planting.
R. solanacearum inoculum consisted of a bacterial suspension containing
109cfu/ml in sterile distilled water. Inoculation was done within
two hours of inoculum preparation using the soil drenching and root-severing
method described by Winstead and Kelman (1952). Treatments were arranged in
a completely randomised design with three replicates. Plants were rated on
a weekly basis for bacterial wilt severity up to 30 days post inoculation.
A scale of 0-5 (from wilted leaves to death) where, 0 = no symptoms, 1 = 1
leaf wilted, 2 = 2 or 3 leaves wilted, 3 = all the leaves wilted except the
top 2 or 3 leaves, 4 = all leaves wilted, 5 = plant dead, was used (Winstead
and Kelman, 1952). The experiment was repeated to validate the results. Data
were subjected to analysis of variance and means were separated using Fisher's
protected least difference test at P=0.05.
RESULTS
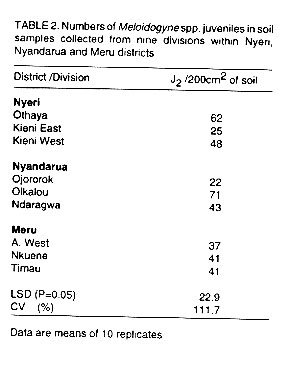
Population density of Meloidogyne spp. in potato fields. Table
1 shows the second stage juveniles of Meloidogyne spp. in soil
obtained from different altitude ranges. The correlation between altitude
and population densities of Meloidogyne spp. juveniles were not significant
(P>0.05). The mean population densities of Meloidogyne spp. juveniles
in soil collected from Nyeri, Nyandarua and Meru districts are depicted in
Table 2. The mean population of Meloidogyne
juveniles in 200cm3 soil was 45 in Nyeri and Nyandarua districts
and 31 in Meru district. Variations in juvenile populations of Meloidogyne
spp. were significant (P<0.05) among divisions of the districts.
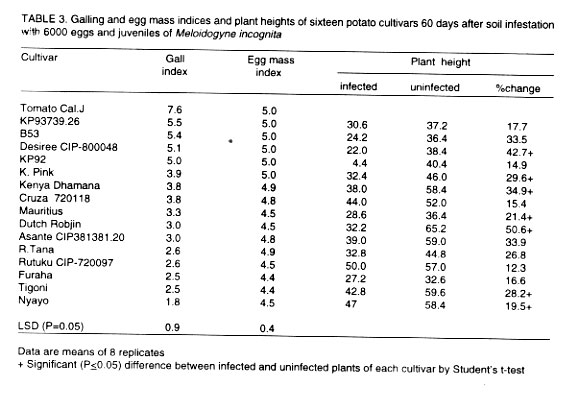
Reaction of potato cultivars to Meloidogyyne incognita. Differences
in galling and egg mass indices were significantly (P=0.05) different among
the potato cultivars, 60 days after soil infestation with 6000 eggs and juveniles
of M. incognita (Table 3). None
of the cultivars was immune to M. incognita. Galling index was highest
(5.5) in cv. KP93739.26 and lowest (1.9) in cv. Nyayo. The susceptible control,
tomato cv. Cal J, had the highest (7.6) galling index. With the exception
of potato cv. KP93739.26, tuber infection was not observed.
Differences in nematode reproduction among the cultivars tested were highly
significant (P < 0.01). All cultivars supported nematode reproduction
with highest (5.0) egg mass index being recorded in cultivars Kerr's
Pink, KP93739.26, Desiree (CIP-800048), KP92633.26, B53 and the susceptible
control, tomato cv. Cal J. Nematode reproduction was lowest in cultivars Nyayo
and Tigoni (CIP-381381.13) both of which had an EMI of 4.4. A positive and
significant (r = 0.484; P=0.001) correlation existed between egg mass index
and galling index.
Nematode infection caused reduction of plant height in all the potato cultivars
screened (Table 3). Cultivar Rutuku CIP-720097
recorded the lowest (12.4%) reduction while the highest (50.6%) reduction
was observed in cultivar Dutch Robjin. Differences in plant height between
infected and uninfected plants were significant (P=0.05) in cultivars Desiree,
K.Pink, K. Dhamana, Mauritius, Tigoni and Nyayo.
Effect of Meloidogyne incognita on development and severity of bacterial
wilt of potato. Differences in bacterial wilt severity were significant
(P<0.05) among the treatments for all the three cultivars used in
this test (Table 4). Bacterial wilt severity
was higher in plants inoculated with R. solanacearum and M. incognita
than those inoculated with R. solanacearum alone.
Analysis of variance revealed significant (P<0.05) differences
in bacterial wilt severity among the three cultivars. Plants whose roots were
severed and inoculated with the two pathogens had the highest (3.4) overall
bacterial wilt severity. Wilt severity was lower (3.2) but not significantly
(P>0.05) different in plants inoculated with both pathogens without root
severing. Wilt severity in plants inoculated with R. solanacearum without
root-wounding was significantly (P=0.05) lower (1.1) than in plants inoculated
with R. solanacearum and whose roots were severed (2.6).
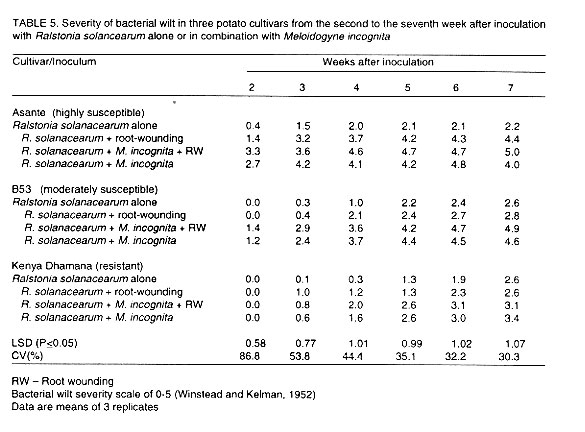
Disease progress in the three potato cultivars is depicted in Table
5. For all the cultivars, significant (P=0.05) differences in bacterial
wilt severity were observed among potato plants inoculated with R. solanacearum
alone without root wounding and those inoculated with R. solanacearum
and M. incognita throughout the seven weeks. Bacterial wilt
severity was consistently higher in plants whose roots were severed. When
plants inoculated with R. solanacearum alone were compared with those
inoculated with R. solanacearum in combination with M. incognita,
significant (P<0.05), differences in wilt severity were observed
in each of the three cultivars throughout the assessment period (Table
5). Bacterial wilt development was faster and the severity consistently
higher in plants inoculated with the two pathogens. Similarly, comparisons
between plants inoculated with the two pathogens simultaneously with severed
roots and those treated the same way but with unsevered roots indicated non-significant
(P=0.05) differences in each cultivar and for every week (Table
5).
Bacterial wilt severity was higher and development faster in wounded plants
inoculated with R. solancearum in combination with M. incognita.
Likewise bacterial wilt severity was higher in potato plants inoculated with
R. solanacearum with root-wounding than in plants inoculated with both
R. solanacearum and M. incognita without root wounding.
DISCUSSION
The occurrence of Meloidogyne spp. juveniles in soil samples obtained
from potato fields in Nyeri, Meru and Nyandarua districts supports a previous
report that root-knot nematodes are commonly associated with potato in the
tropics (Jatala and Bridge, 1990). Although temperature is known to influence
all components of nematode life processes and host parasite relations (Noe
and Sikora, 1990), the differences in root-knot nematode populations recovered
from the potato fields could not be attributed to variation in temperature.
The low populations of Meloidogyne spp. in Meru district was unexpected
since the district experiences the highest temperature. However, apart from
temperature, other factors such as moisture and aeration may have come into
play in determining the population dynamics of Meloidogyne spp. (Sasser
and Freckman, 1987).
Root galling, nematode reproduction and stunted growth were observed on
all nematode-infected potato cultivars with no indications of immunity to
M. incognita. Potato cultivars, however, were differentially stunted,
confirming reports of differential varietal response to nematode infestation
(Jatala and Bridge, 1990). A compatible reaction characterized by penetration
and development of M. incognita larvae to maturity was observed in
all the cultivars. Some cultivars such as KP93739.26, B53, and KP 92633.26
had high gall index and egg mass index but were not significantly stunted
following nematode infection. Such cultivars may be tolerant to nematode infection.
Tubers obtained from nematode infected plants did not show galls or egg masses.
According to Jatala (1975), tubers become infected when conditions are optimal.
Penetration of M. incognita juveniles and symptom development in tubers
require relatively higher temperatures (Charchar and Moita, 1996) than those
required for parasitism of potato roots (Mai et al., 1981). Lack of
galling on tubers, 60 days after inoculation in all the potato cultivars except
KP 93739.26, may be attributed to the low temperatures that prevailed during
the experiment. The mean atmospheric temperature at the time of the experiment
was 17.20C. Although no cultivar showed immunity to Meloidogyne,
cultivars such as Nyayo, Tigoni, and Furaha can be recommended for soils heavily
infested with Meloidogyne spp.
Although results from greenhouse screening may not be reproduced under field
conditions (Janssen et al., 1996), it is conceivable that damage as
a result of Meloidogyne infection in the field would be more severe
because nematode infected roots are predisposed to other pathogens (Jatala
and Bridge, 1990). In this evaluation, other soil-borne pathogens were destroyed
through sterilization of the potting mixture. Effects of nematode damage,
which are usually exaggerated by moisture stress, due to inability of roots
to absorb moisture effectively, may have been reduced by regular watering
of the plants.
Greenhouse studies revealed that infection by R. solanacearum in the
presence of root-knot nematodes resulted in a higher severity and a faster
development of bacterial wilt. Similar findings were reported by Ravichandra
et al. (1990) and Akiew et al. (1991). The fact that bacterial
wilt development was faster and severity consistently higher in plants inoculated
with R. solanacearum in combination with Meloidogyne incognita than
in plants inoculated with R. solanacearum alone suggests that the resistance
mechanism(s) of potato plants may have been weakened in the presence of Meloidogyne.
Nagesh et al. (1997) suggested a breakdown of resistance in tomato
plants inoculated with R. solanacearum together with M. incognita.
Our results showed that bacterial wilt severity was lower in potato cv.
B53 plants inoculated with R. solanacearum with root-wounding than
in plants inoculated with R. solanacearum and M. incognita without
root wounding. This shows that the role of M. incognita in bacterial
wilt development is more than just providing avenues for the entry of bacterium.
According to Napiere and Quimio (1980) and Samuel and Mathew (1983), the synergism
between the two pathogens was mostly associated with wounds caused during
penetration of larvae into roots. However, findings by Sitaramaiah and Sinha
(1985) suggested that nematode induced stress was more important as a wilt-triggering
factor than the wounding. Biochemical and physiological changes resulting
from infection by root-knot nematodes may be responsible for the enhancement
of wilt. According to Trudgill (1991), the biochemistry of a plant is considerably
altered following nematode infection. These changes may either weaken the
chemical resistance mechanisms of the plant or modify conditions in the infected
potato tissue making the plant more suitable for bacterial colonization (Khan,
1993). This is in harmony with the suggestion by Jatala (1975) that infection
by one pathogen may alter the response of a host to subsequent infection by
another.
The result of this study reinforces the need to control root-knot in fields
heavily infested with R. solanacearum in order to gain maximum benefit
from the use of wilt resistant potato cultivars. Use of potato cultivars resistant
to Meloidogyne are recommended as a component of integrated bacterial
wilt disease management.
REFERENCES
-
Ajanga, S. 1993. Status of bacterial wilt in Kenya. In:
Bacterial wilt. Hartman, G. L and Hayward, A. C. (Eds.), pp. 338-340.
ACIAR Proceedings No. 45, Canberra Australia
-
Akiew, E., Trevorrow, P. R. and Waite, C. A. 1991. Tobacco
bacterial wilt investigations in North Queensland. ACIAR Bacterial Wilt
Newsletter 8:11-12.
-
Bird, G.W. 1981. Management of plant nematodes in potato
production. In: Advances in potato pest management.
-
Lashomb, J. H. and Casogrande, R. (Eds.), pp. 223-244.
-
Brown, C.R., Mojtaahedi, H. and Santo, G.S. 1989. Comparison
of reproductive efficiency of Meloidogyne chitwoodi on Solanum
bulbocastanum in soil and in vitro tests. Plant Disease 73:957-
959
-
Charchar, J.M. and Moita, A.W. 1996. Reacao de cultivares
de batat a infeccao por Meloidogyne incognita raca 1. Horticultura
- Brasileira 14:189-193.
-
Dropkin, V.H. 1980. Introduction to plant nematology.
Second edition. John Wiley & Sons, Inc. USA. 304 pp.
-
Evans, K. and Trudgill, D. L. 1992. Pest aspects of potato
production. In: The Nematode Pests of the Potato Crop.
-
Harns, P. M. (Ed.). Chapman Hall Press, London.
-
FAO, 1995. Potatoes in the 1990s. Situation and prospects
of the world potato economy. CIP, Lima, Peru.
-
Haider, M.G., Nath, R.P., Thakur, S.C. and Ojha, K.L. 1987.
Interaction of Meloidogyne incognita and Pseudomonas solanacearum
on tomato plants. Indian Journal of Nematology 17:174 -176.
-
Hayward, A.C. and Hartman, G.L. 1994. Bacterial wilt: The
disease its causative agent Pseudomonas solanacearum. CAB International,
Wallingford, UK.
-
Hooper, D.J. 1990. Extraction and processing of plant and
soil nematodes. In: Plant parasitic Nematodes in subtropical and tropical
agriculture. Luc, M., Sikora, R.A. and Bridge, J. (Eds.), pp. 45-68.
CAB International, Wallingford.
-
Janssen, G. J.W., Janssen, R., Novel-A-van, B., Verkerk-Bakker
and Hoogendoorn, J. 1996. Expression of resistance to the root-knot nematodes,
Meloidogyne hapla and M. fallax in wild Solanum spp.
under field conditions. European
-
Journal of Plant Pathology 102: 859-865.
Jatala, P. 1975. Root-knot nematodes (Meloidogyne spp.) and their
effect on potato quality. In: Proceedings of the 6 th
-
Triennial Conference of the EAPR. 15-19 September
1995, Wageningen, Netherlands. 194 pp.
-
Jatala P. and Bridge, J. 1990. Nematode parasites of root
and tuber crops. In: Plant Parasitic Nematodes in Sub-tropical and Tropical
Agriculture. Luc, M., Sikora, R A. and Bridge, J. (Eds.), pp. 137-149.
CAB International, Willingford UK.
-
Johnson, H. A. and Powell, V. T. 1969. Influence of root-knot
nematodes on bacterial wilt development in flue-cured tobacco. Phytopatholology
59:486-91.
-
Khan, M.W. 1993. Mechanisms of interactions between nematodes
and other plant pathogens. In: Nematode Interactions. Khan, M.W.
(Ed.), pp. 55-78. Chapman and Hall. Madras, India.
-
Mai, W.F., Brodie, B.B., Harrison, M.B. and Jatala, P. 1981.
Nematodes. In: Compedium of potato diseases. Hooker, (Ed.), pp. 93-101.
American Phytopathological Society, St. Paul, MN.
-
Michieka, A.O. 1993. Screening CIP potato germplasm for
resistance to Pseudomonas solanacearum E.F. Smith. In:
-
Proceedings of a Workshop on bacterial wilt of potato
caused by Pseudomonas solanacearum. Bujumbura, Burundi, February 22-26,
1993. pp. 27-31.
-
Nagesh, M., Chakrabarti, S.K., Shekhawat, G.S. and Gedewar,
A.V. 1997. Evaluation of potato accessions for their combined resistance
to Pseudomonas solanacearum and Meloido-gyne incognita. Pest
Management in Horticultural Ecosystems 3:17-20.
-
Napiere, C.M. and Quimio, A.J. 1980. Influence of root-knot
nematode on bacterial wilt severity on tomato. Annals of Tropical Research
2:29 - 39.
-
Noe, J. P. and Sikora, R.A. 1990. Effects of tropical climate
on the distribution and host-parasite relationship of plant parasitic nematodes.
In: Plant parasitic nematodes in sub tropical and tropical agriculture.
Bridge, J., Sikora, R.A. and Luc, M. (Eds.), pp. 558-583. CAB1, Willinfford,
UK.
-
Omwega, C.O., Thomason, I.J. and Roberts, P.A. 1988. A non-
destructive technique for screening bean germplasm for resistance to Meloidogyne
incognita. Plant Disease 72:970-72.
-
Ravichandra, N. G., Krishnappa, K. and Setty, K.G.H. 1990.
Interaction of Meloidogyne javanica and race 1, race 2, race 3 of
Meloidogyne incognita with Pseudomonas solanacearum
on a few Brinjal lines. Indian Journal of Nematology 20:138-147.
-
Samuel, M. and Mathew, J. 1983. Role and association of
root-knot nematode Meloidogyne incognita in the introduction
of bacterial wilt of ginger incited by Pseudomonas solanacearum.
Indian Phytopathology 36:398 - 399.
-
Sasser, J.N. and Freckman, D. W. 1987. A world perspective
on nematology: The role of the society. In: Vistas on nematology.
Veech, J.A. and Dickson, D.W. (Eds.), pp. 7- 14. Society of Nematologists.
-
Sequeira, L. 1992. Bacterial wilt: Past, present and future.
In: Bacterial wilt. Hayward, A.C. and Hartman, G.L. (Eds.).
-
Proceedings of an international conference held at Kaoshiung,
Taiwan, 28-31 October, 1997.
-
Sijmons, P.C., Alkinson, H.J. and Wyss, U. 1994. Parasitic
strategies of root-knot nematode and associated cell responses. Annual
Review Phytopathology 32:235-59.
-
Sitaramaiah, K. and Sinha, S.K. 1985. Histological aspects
of Pseudomonas solanacearum and root- knot nemotode complex in brinjal.
Indian Journal of Nematology 14:175 - 178.
-
Taylor, A. L. and Sasser, J. N. 1978. Biology, identification
and control of root-knot rematodes.International Meloidogyne Project.
North Carolina State University Graphics, 111 pp.
-
Trudgill, D.L. 1991. Resistance to and tolerance of plant
parasitic nematodes in plants. Annu. Rev. Phytopathology 29:167-192.
-
Winstead, N.N. and Kelman, A. 1952. Inoculation techniques
for evaluating resistance to Pseudomonas solanacearum. Phytopathology
42:628-34.
-
Zech, W.M. 1971. A rating scheme for field evaluation of
root-knot nematode infestations. Pflanzenschutz-Nachrichten. Bayer AG
24: 141-144.
© Copyright 2001, African Crop Science Society
The following images related to this document are available:
Photo images
[cs01072t5.jpg]
[cs01072t1.jpg]
[cs01072t3.jpg]
[cs01072t2.jpg]
[cs01072t4.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}