 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
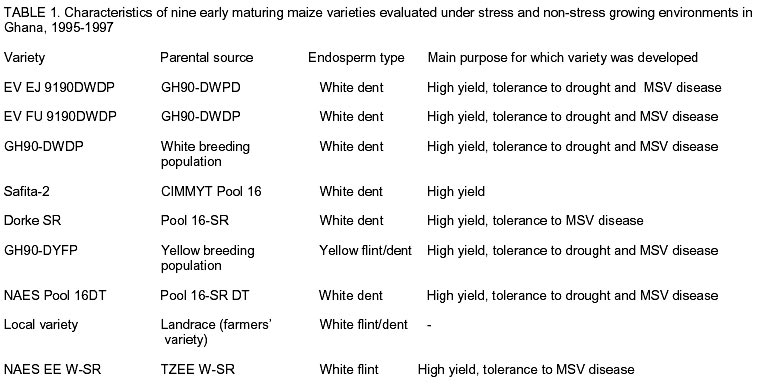
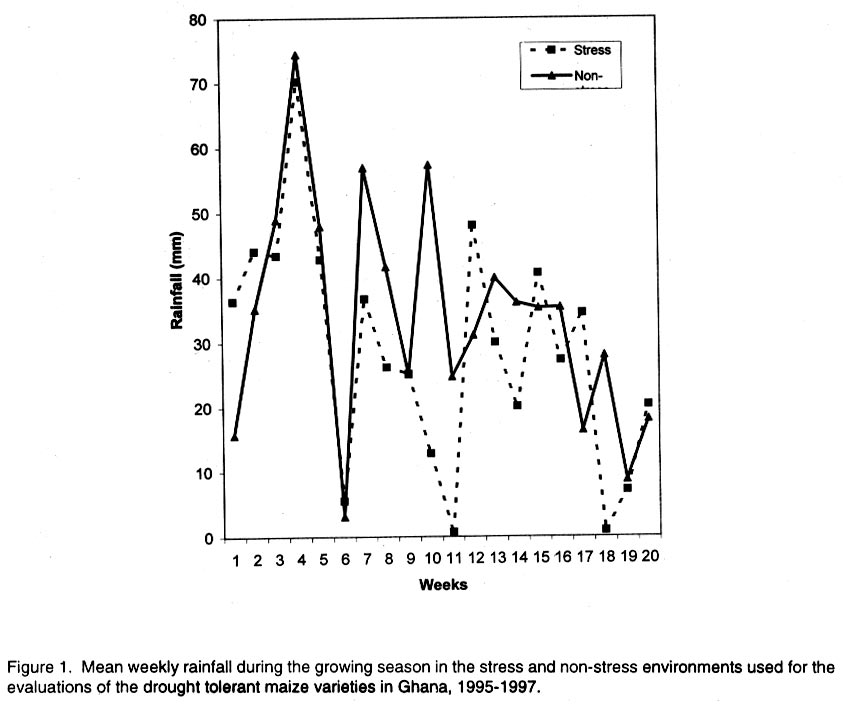
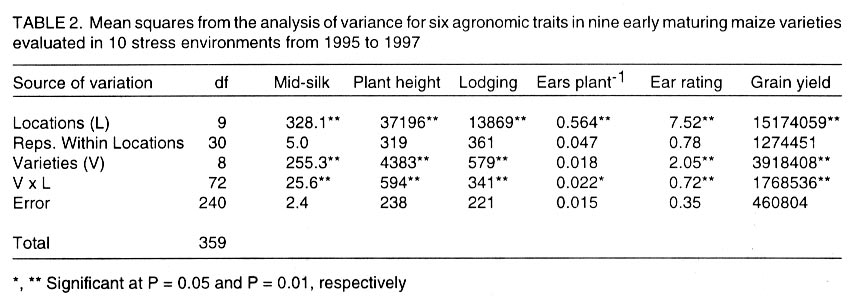
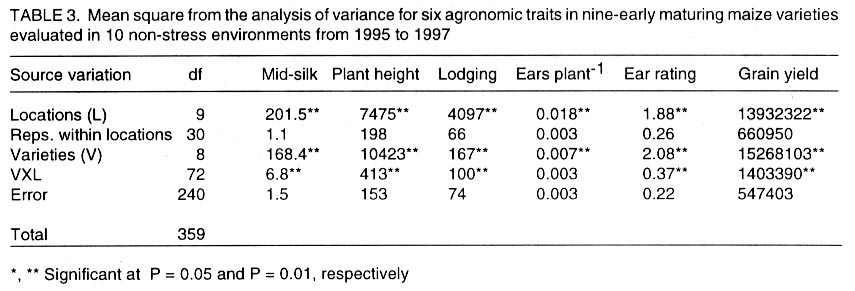
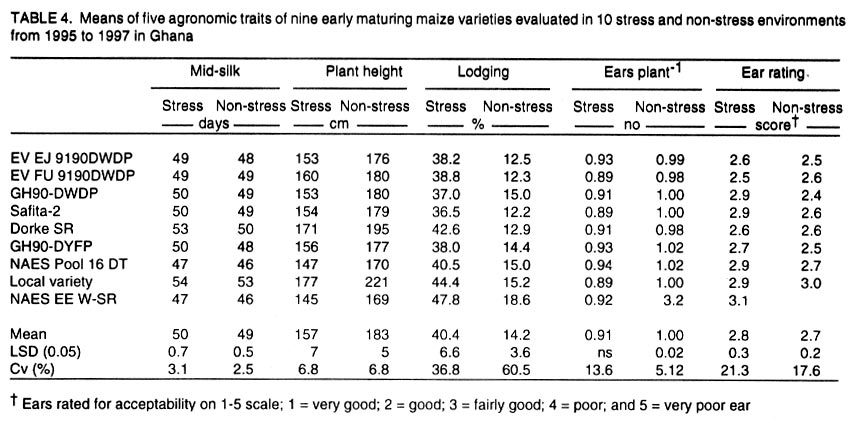
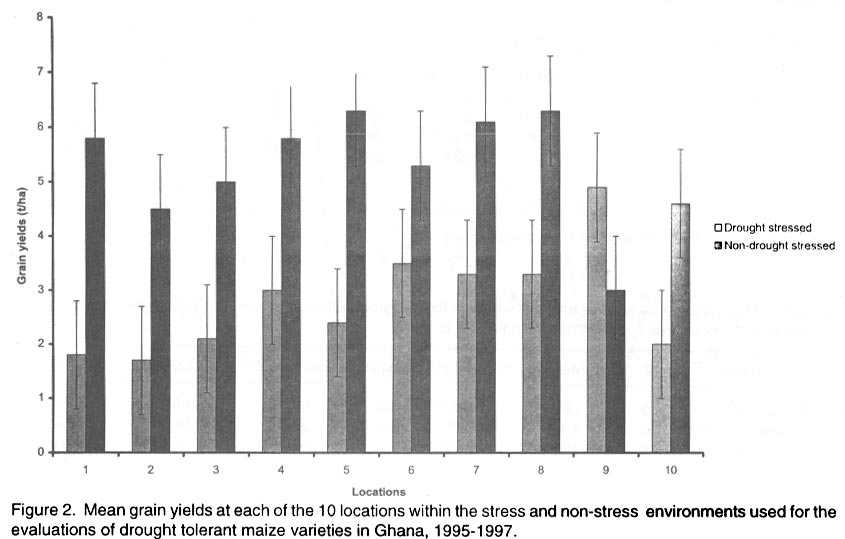
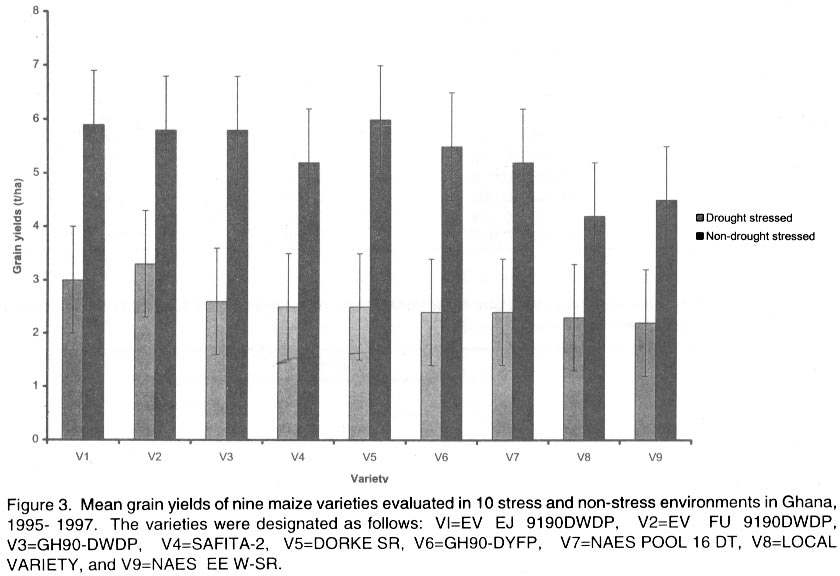
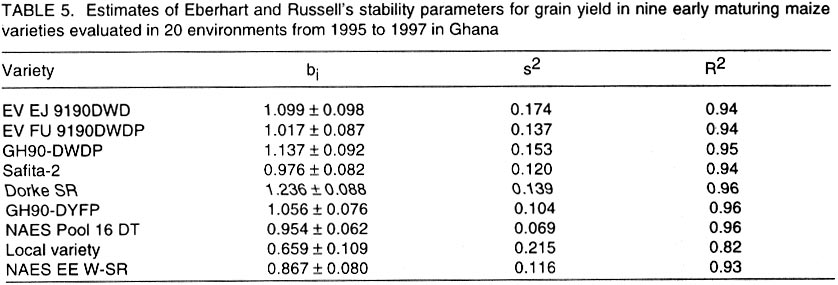
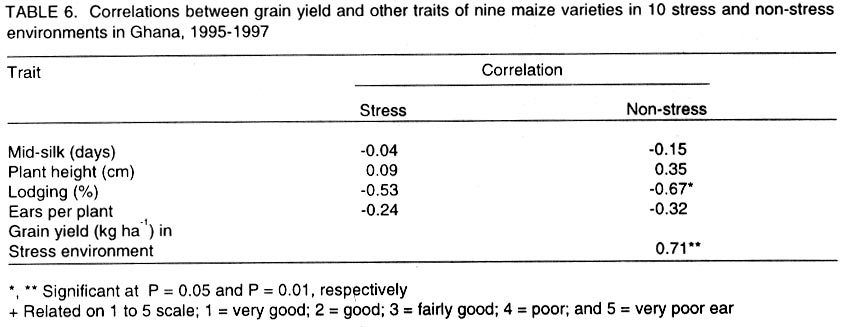
African Crop Science Journal, Vol. 10. No. 1, 2002 pp. 1-9 POTENTIAL OF ELITE MAIZE COMPOSITES FOR DROUGHT TOLERANCE IN STRESS AND NON-drought ENVIRONMENTS P.Y.K. SALLAH, K. OBENG-ANTWI and M.B. EWOOL Crops Research Institute, CSIR, P. O. Box 3785, Kumasi, Ghana (Received 11 December, 1999; accepted 11 November, 2001) Code Number: cs02001 ABSTRACT Drought stress is a major factor limiting the productivity of maize (Zea mays L.) in West and Central Africa. This study was conducted to assess drought tolerance/susceptibility in nine early maturing maize composites, including five drought tolerant selections. The varieties were evaluated in 5 drought-stressed and 5 non-drought stressed environments in Ghana, West Africa, from 1995 to 1997. Effects due to environment (E), genotype (G) and G x E interaction were highly significant (P<0.01) for grain yield, 50% silk emergence, plant height, lodging, ears per plant, and ear rating in both environments (drought and non-drought stressed). In the stress environment, grain yields of the varieties ranged from 2.21 to 3.12 t ha-1, while in the favourable environment yields ranged from 4.17 to 5.96 t ha-1. Two drought tolerant selections (EVEJ 9190DWDP & EVFU 9190DWPD) out-yielded Dorke SR (an improved check) by 16.0% and the Local (landrace) variety by 30.8% in the stress environment. In the favourable environment, grain yields were similar, averaging 5.85 t ha-1 for the two elite varieties and the improved check. Grain yield in the stress environment was positively associated (r = 0.71, P<0.01) with yield in the non-stress environment. Estimates of Eberhart and Russell's stability parameters across environments were bi = 1.04 (s2=0.13) for the improved varieties and bi=0.65 (s2=0.22) for the local variety. Key Words: Ghana, grain yields, productivity, selections, Zea mays Résumé La sécheresse est le plus grand facteur limitant la production de mais (Zea mays L.) en Afrique de l'ouest et centrale. Cette étude était conduite pour estimer la tolérance de neuf jeunes plantes à la sécheresse. Cinq de neuf sélections sont reputées tolérantes à la sécheresse. Les variétés étaient évaluées dans cinq environnement de sécheresse permenante et non permenante du Ghana de 1995- 1997. Les effets due à l'environnement (E); génotype (G) et à l'intéraction GxE étaient significativement élévés (P<0.01) pour le rendement en grains (50% 'silk' emergence) , taille de la plante, spathes, les épis par plante, et le taux d'épis dans les deux environnements. Dans l'environnement stressé les rendements en grains ont rangé entre 2,21 et 3,12 t ha-1; et dans les conditions favorables ils ont rangé entre 4,2 et 5,96 t ha-1. Deux sélections tolérantes à la sécheresse (EVEJ 9190 DWDP et EVFV 9190 DWPD) ont surpassé le Dorke SR (Variété améliorée) par 16% et la variété locale (Landrace) par 30.8% dans un environnement stressé. Dans les conditions favorables, les rendements en grains étaient similaires avec une moyenne de 5.87 t ha-1, pour deux variétés élites et variétés améliorées. Le rendement en grains dans l'environnement stressé était positivement associé (R= 0.71, P< 0.01) à celui de l'environnement non stressé. Une estimation des paramètres de stabilité de Eberhart et Russel à travers environnements étaient de bi=1,04 (S2 = 0.13) pour les variétés améliorées et bi = 0,65 (S2= 0.22) pour la variété locale. Mots Clés: Ghana, rendement en grain, productivité, sélections, Zea mays INTRODUCTION Maize is an important staple crop in rain-fed production systems of West and Central Africa. Though total maize production increased tremendously in the region over the past three decades, productivity per unit area remains low. For example, in Ghana, maize production increased from 481,600 t in 1970 to 1,034,200 t in 1997 (PPMED, 1991, 1998). During the same period, average maize yields increased from 1.07 to 1.55 t ha-1 (PPMED, 1991, 1998). The low productivity of maize in the region has been attributed to two main abiotic stresses, low soil fertility and drought. Of the two, drought is perhaps more difficult to manage by the small-scale farmers who are the major maize producers in this region. This is because the occurrence of drought is unpredictable. Thus, varieties suitable for farmers in this zone should perform well under both drought-stressed and favourable conditions. Maize is extremely susceptible to drought stress at flowering. Stress occurring on a single day at this stage results in twice as much grain yield reduction as any other stage (Shaw, 1976; Grant et al., 1989; NeSmith and Ritchie, 1992). The extreme susceptibility of maize to drought stress has been attributed to the spatial separation of male and female flowers on the same plant, differential partitioning of assimilates in favour of the tassel versus the ear when the plants are stressed at flowering (Edmeades et al., 1993), and the negative effects of water deficits on hormonal and enzymatic activities in the newly-fertilised ovary (Westgate, 1997). Drought at flowering is reported to reduce flux of assimilates, especially sucrose, to developing kernels, resulting in abortion of fertilised kernels shortly after fertilisation (Schussler and Westgate, 1995; Zinselmeier et al., 1995a, b). Hence, variation in grain yield under drought stress seems to be due mainly to variation in number of kernels per plant. Selection to improve kernel number and its associated traits under drought stress, therefore, resulted in improvement of grain yield under stress (Edmeades et al., 1997). Several selection strategies for improving the productivity of maize under drought stress have been investigated but the results are inconclusive. These include selection only under favourable conditions (Arboleda-Rivera and Compton, 1974; Johson and Geadelmann, 1989; Martinez-Barajas et al., 1992), selection only under drought stress (Arboleda-Rivera and Compton, 1974), selection under both favourable and stress conditions involving multilocation testing (Arboleda-Rivera and Compton, 1974), and selection under drought stress at one location together with multi-location testing (Byrne et al., 1995). Johnson and Geadelmann (1989) found that progress from selection for yield under irrigation equalled that from selection under drought stress when evaluated under drought stress, but was greater when evaluated under favourable conditions. However, Arboleda-Rivera and Compton (1974) and Martinez-Barajas et al. (1992) reported that grain yields from selection under favourable conditions were greatly reduced under drought stress. Arboleda-Rivera and Compton (1974) also reported that: (1) selection only under drought stress resulted in increased yield under stressed and un-stressed conditions, and (2) selection in a combination of stressed and un-stressed environment resulted in a lower gain per cycle than when selection was done in either stressed or favourable environments. In the early 1980s, breeders at the Crops Research Institute, Kumasi, Ghana, initiated a breeding programme to improve tolerance to drought stress in early maturing maize populations through cyclical selection in alternating favourable and stressed environments. These efforts resulted in the development of two breeding populations and experimental varieties from the improved cycle of the populations. However, the performance of these varieties under varying conditions has not been evaluated. The objectives of this study therefore were to (i) determine the productivity of these varieties under favourable and drought conditions prevailing in the major maize producing zones of Ghana, (ii) determine the relationship between varietal performance under favourable and stress conditions, and (iii) identify high and stable yielding varieties across both environments. Materials and methods Nine early maturing maize varieties, including the farmers' (Local or landrace) variety, were evaluated in the major season at Pokuase and Ohawu (coastal savanna, fine sandy-loam Dystrochrept); Fumesua and Kwadaso (forest, coarse sandy-loam Paleustult); Ejura and Kpeve (transition, fine-coarse sandy-loam, Oxisol); Damongo, Nyankpala and Wa (Guinea savanna, fine sandy-loam Alfisol) and Manga (Sudan savanna, fine sandy-loam Alfisol) from 1995 to 1997. The source materials and other attributes of the varieties studied are presented in Table 1. Zero-tillage was practiced in the fields in the coastal savanna, forest and transition zones, and this comprised application of Glyphosate (Round-up) at 1.5 kg a.i. ha-1 two weeks before planting the trials. In the Guinea and Sudan Savanna zones, the fields were ploughed, harrowed and ridged before planting. A randomised complete block design with 4 replicates was used at each site. A plot consisted of four 5-m rows of each variety at 66,000 plants ha-1 density in the interior savanna locations and at 62,000 plants ha-1 in the coastal savanna, forest, and transition zones. Pre-emergence chemical weed control was used and this consisted of application of a combination of Pendimethalin and Gesaprim at 1.5 kg a.i.ha-1 and 1.0 kg a.i ha-1, respectively at planting. Where there was lush vegetation prior to planting, Paraquat was applied at 1.0 kg a.i.ha-1 in addition to Pendimethalin and Gesaprim. Hand weeding was also done as a follow-up when necessary to keep the plots free of weeds. Fertilisation was by spot-application of 45 kg N ha-1 and 45 kg P2O5 ha-1 at 8-10 days after planting at all sites. Additional 45 kg N ha-1 was applied as side-dressing using urea or sulphate of ammonia at 3-4 weeks after planting. Data were recorded from the two middle rows of the plot of each variety on grain yield at 15% moisture, days to 50% silk emergence, plant height, total lodging, number of ears per plant, and ear acceptability rating. Ear acceptability was rated on a 1-5 scale, where 1 = very good; 2 = good; 3 = fairly good; 4 =poor and 5 = very poor ear. The weather pattern at each site was monitored for periods of drought, particularly at the flowering stages of the crop. Rainfall data were also recorded at each site. The data were then used to group the sites into a stressed set and a non-stress (favourable) set. Useable agronomic data were obtained from 10 sites in the three years for each environment. The data were analysed by site (location) and combined over locations within each set (Steel and Torrie, 1980). Phenotypic correlation coefficients were calculated between grain yields in the stress and non-stress environments and between yield and other agronomic traits for each environment. Grain yield stability in the varieties was assessed using the procedure suggested by Eberhart and Russell (1966). The stability parameters computed from this method were bi and s2, where bi is the linear regression coefficient of the ith variety on the environmental index measured as the mean yield of all the varieties in that environment minus the mean of all the environment, and s2 is the deviation from regression of the ith variety minus the average variance of a variety mean in the jth environment. The hypothesis that a regression coefficient for a variety did not differ from unity, and, the deviation from regression was not different from zero was tested using the method of Eberhart and Russell (1966). RESULTS AND DISCUSSION Mean weekly rainfall during the growing season for the stress and non-stress environments are presented in Figure 1. The values were computed as average of weekly rainfall at the 10 sites, beginning from the week of sowing at each site. Rainfall distribution during the first six weeks after planting were similar for both environments (Fig. 1). Thereafter, the non-stress environment received 79.3% more rainfall than the stress environment up to the eleventh week after sowing. The period of differential rainfall coincided with flowering, pollination, and grain filling stages of the varieties. The summarised analyses of variance (mean square values and P levels), combined over locations, for six agronomic traits are presented in Table 2 for the stress environment and in Table 3 for the non-stress environment. Location effects were highly significant (P<0.01) for number of days to 50% silk emergence, plant height, lodging, number of ears per plant, ear acceptability rating and grain yield in the stress and non-stress environments. The effects due to varieties were also highly significant for all the traits in both environments, except for ears per plant in the stress environment. In the stress environment, variety x location interactions were significant (P=0.05) for ears per plant, but were highly significant (P=0.01) for the other traits (Table 2). These effects were also highly significant (P=0.01) for all the traits, except ears per plant, in the non-stress environment (Table 3). The significant variety x location interactions observed for these traits showed that the varieties responded differentially to the sites within each environment. The significant interaction also revealed differences among sites within each environment. Variations in soil characteristics at the sites probably accounted for these differences. Differences among the varieties for number of days to silking in the stress and non-stress environments were small but significant (Table 4). However, the type of environment did not seem to have appreciable influence on flowering in the varieties. Plant heights differed significantly among the varieties, ranging from 1.5 to 1.7 m in the stress environment and from 1.7 to 2.2 m in the non-stress environment (Table 4). On the average, plants in the stress environment were 14.2% shorter than those in the non-stress environment. In contrast, plants in the stress environment lodged more severely, averaging 26.2% higher than in the non-stress environment (Table 4). Edmeades et al. (1997) also observed high lodging in maize under moisture stress and attributed this phenomenon to dehydration of the stem in the stress environment. There was no significant difference among varieties for number of ears per plant in the stress environment (Table 4). Small but significant differences were, however, detected among the varieties for this trait in the non-stress environment. Significant differences were also observed among varieties for ear acceptability rating in both environments (Table 4). Type of environment had no significant influence on ear acceptability rating. This was expected because ears of each variety were rated relative to the others in each environment, thus eliminating environmental influences. Mean grain yields at each of the 10 locations within the stress and the non-stress environments are presented in Figure 2. Mean yields in the stress and the non-stress environments were 2.59 and 5.34 t ha-1, respectively. Hence, yields in the stress environment were 106% lower than in the non-stress environment (Fig. 2). The yield reduction in the stress environment was attributed to drought during the reproductive stages of the maize crop in this environment. Yields of 1.75 to 3.47 t ha-1 were observed at the 5 drought stress sites and 4.52 to 6.12 t ha-1 at the 5 non-drought stress sites (Fig. 2). Mean yields of the varieties ranged from 2.21 to 3.21 t ha-1 in the stress environment and from 4.23 to 5.96 t ha-1 in the non-stress environment (Fig. 3). Populations and varieties improved for tolerance to drought stress out-yielded the other improved varieties by 11.2% in the stress environment and by 7.4% in the non-stress environment (Fig. 3). Two experimental varieties (EV EJ 9190 DWDP & EV FU 9190 DWPD) significantly (P<0.05) out-yielded all the other varieties in the stress environment. Under stress, these varieties yielded 20% higher than Dorke SR, the improved commercial variety. However, the two varieties gave similar yielded as Dorke SR and GH 90 DWDP in the non-stress environment. The lowest yielding varieties were NAES EE W-SR in the stress environment and the local variety in the non-stress environment. The yield superiority of EV EJ 9190 DWPD and EV FU 9190 DWDP over the other varieties observed under drought stress was attributed to intentional selection for tolerance to this stress in our breeding programme. The two varieties were developed from GH90DWDP which had been improved for seven cycles using half-sib family selection under stress as well as favourable conditions (GGDP, 1991). Dorke SR is the improved check variety currently planted by farmers throughout Ghana. Since the two new varieties out-yielded Dorke SR by as much as 20% under stress but were similar to Dorke SR in the favourable environment, adoption of these varieties by farmers will enhance productivity of maize under stress in farmers' fields. There was, therefore, need to evaluate these varieties in farmers' fields to identify the more stable one for release to farmers. Regression analyses of genotype response to an environmental index are often used by plant breeders to estimate stability of performance across varying environments. Estimates of Eberhart and Russell's (1966) stability parameters (bi) for grain yields in the nine varieties across the 20 environments are presented in Table 5. A stable variety is one with unit regression coefficient (bi = 1.0) and a small deviation from regression (s2 = 0) (Eberhart and Russel, 1966). The bi values ranged from 0.87 to 1.24 for the improved varieties and were all not significantly (P<0.05) different from unity. The deviations from linear regression ranged from 0.07 to 0.17 for the improved varieties, and these were all not different from zero. For the local variety, the bi value (0.66) was significantly (P<0.05) less than unity. The deviation from regression (0.22), though the highest, did not also differ significantly from zero. The data showed that all the improved varieties exhibited stable performance whereas the local (landrace) variety exhibited low yield potential across these diverse environments. Phenotypic correlation coefficients between grain yield and five other parameters in the stress and non-stress environments are presented in Table 6. Ear acceptability rating showed strong negative association (P<0.01) with grain yields in both environments. The association of yield with lodging in the non-stress environment was significant. The correlation between yield in the stress environment with that in the non-stress environment was positive and highly significant. In both environments, days to mid-silk, lodging, ears per plant and ear rating were negatively correlated with yield, whereas plant height and ears per plant showed the least relationship with yield. These correlations showed that stand ability (reduced lodging) and ear acceptability rating are important selection criteria for enhancing productivity of maize under moderate levels of moisture stress, as well as in favourable environments. The strong positive association of grain yield in the non-stress environment with yield in the stress environment suggests that a variety that was out-standing in the stress environment was also high-yielding in the non-stress environment. Again, the breeding methodology used for improving the populations and the varieties effectively enhanced productivity in these materials in both types of environments. Results from the study reveal that (1) intermediate level of moisture stress occurred at the 10 sites in the stress environment resulting in severe yield depression in this environment, (2) two drought tolerant composites were more productive than the improved check variety under drought stress but gave similar yields as the improved check in the favourable environment, (3) improved maize varieties were more stable than the landrace variety, and (4) selection in alternating favourable and stressed environments enhanced the productivity of maize in both stress and non-stress environments. REFERENCES
©2002, African Crop Science Society The following images related to this document are available:Photo images[cs02001t5.jpg] [cs02001f3.jpg] [cs02001f2.jpg] [cs02001t1.jpg] [cs02001t2.jpg] [cs02001f1.jpg] [cs02001t3.jpg] [cs02001t4.jpg] [cs02001t6.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}