 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 10. No. 1, 2002, pp. 51-66 SUGARBEET LEAF GROWTH AND YIELD RESPONSE TO SOIL WATER DEFICIT Y.A. Abayomi andWright1 Department of Crop Production, University of llorin, P.M.B. 1515, llorin,
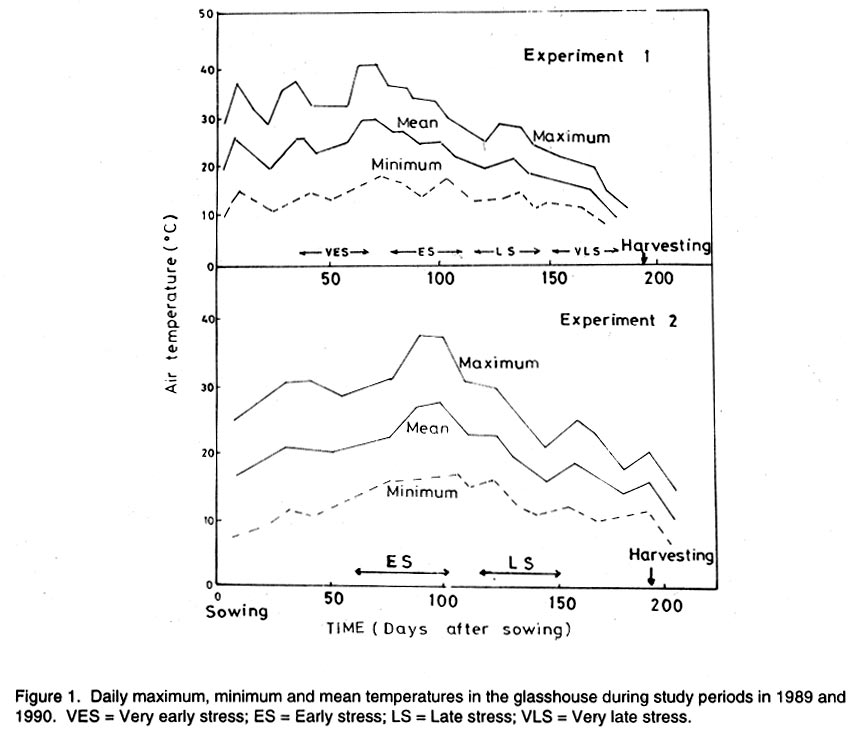
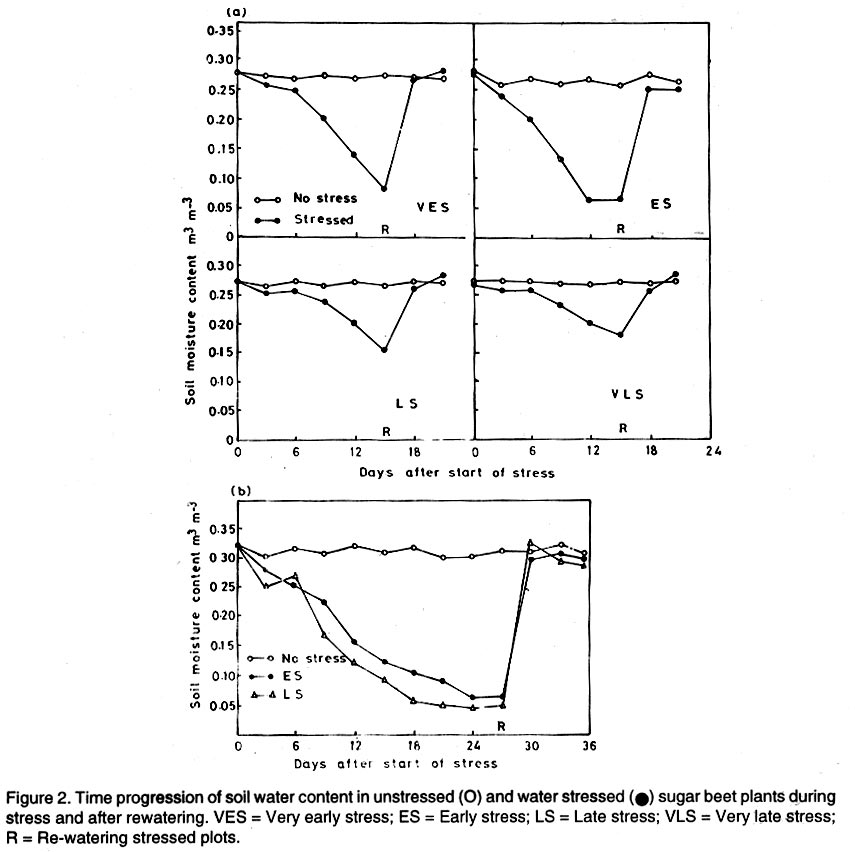
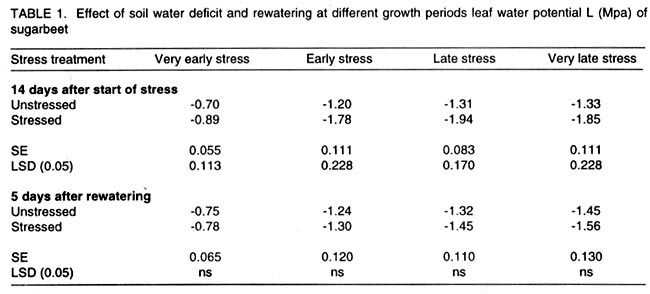
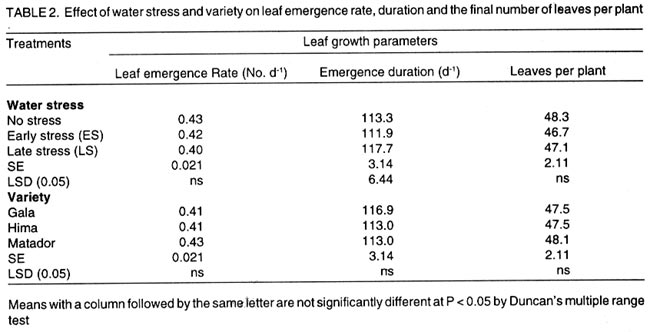
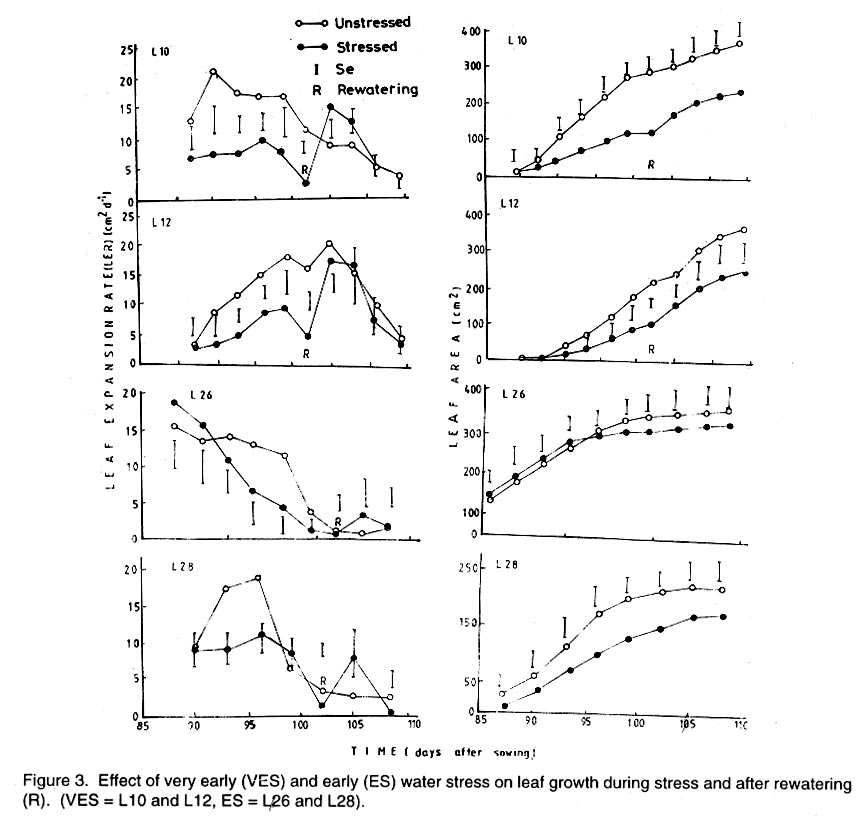
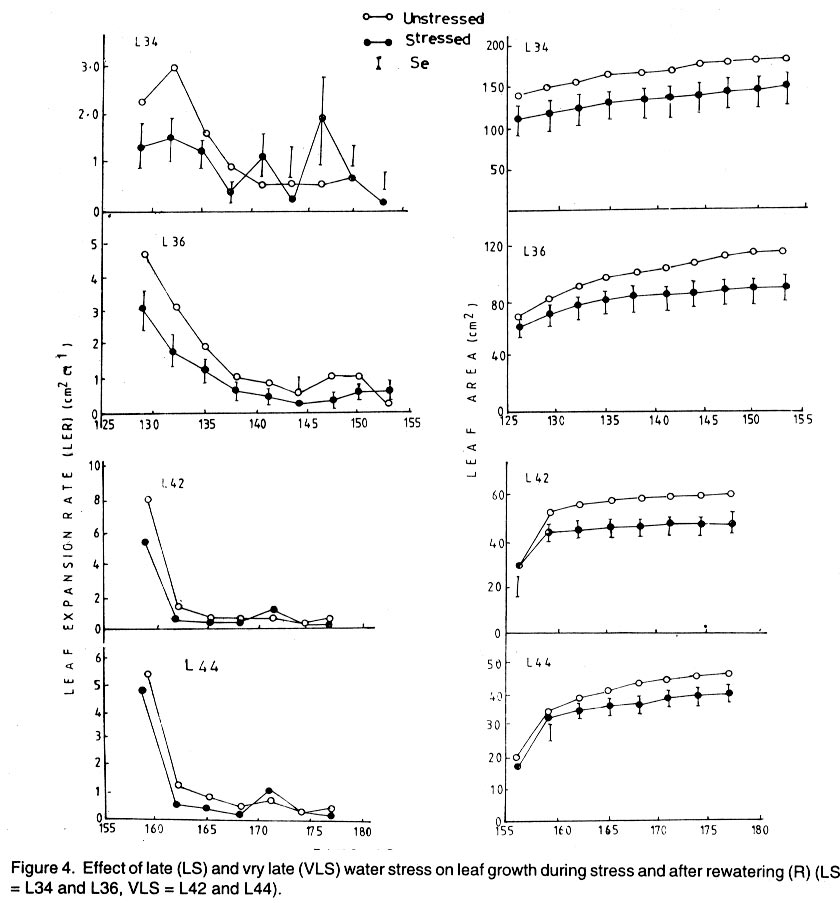
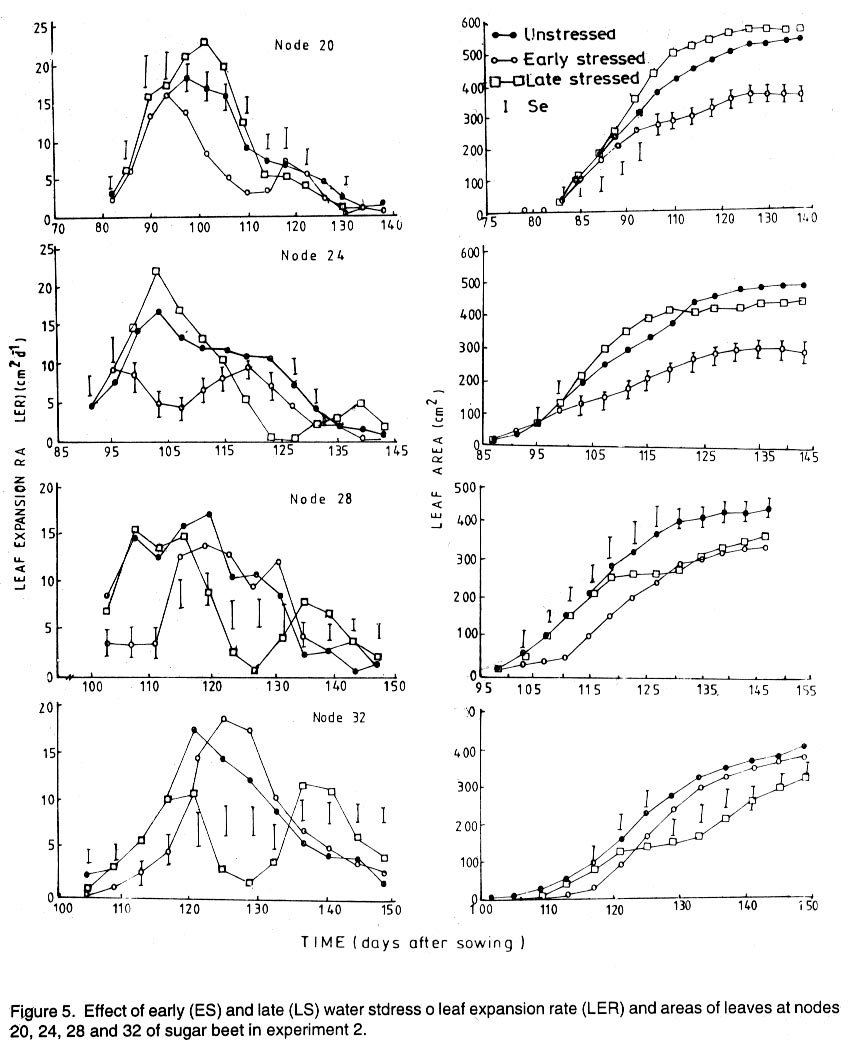
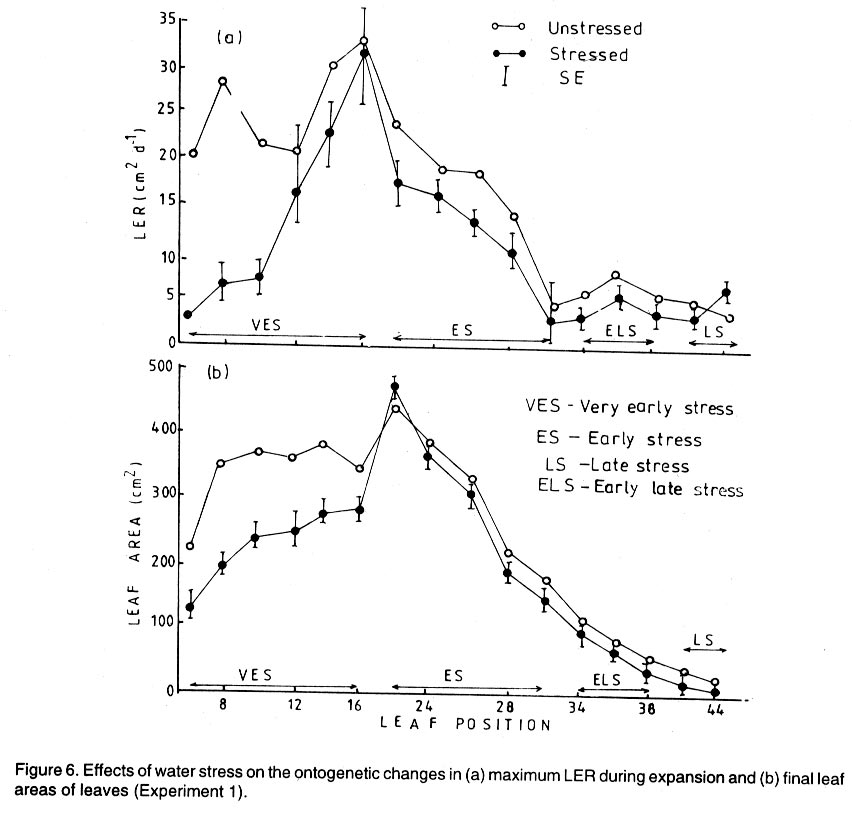
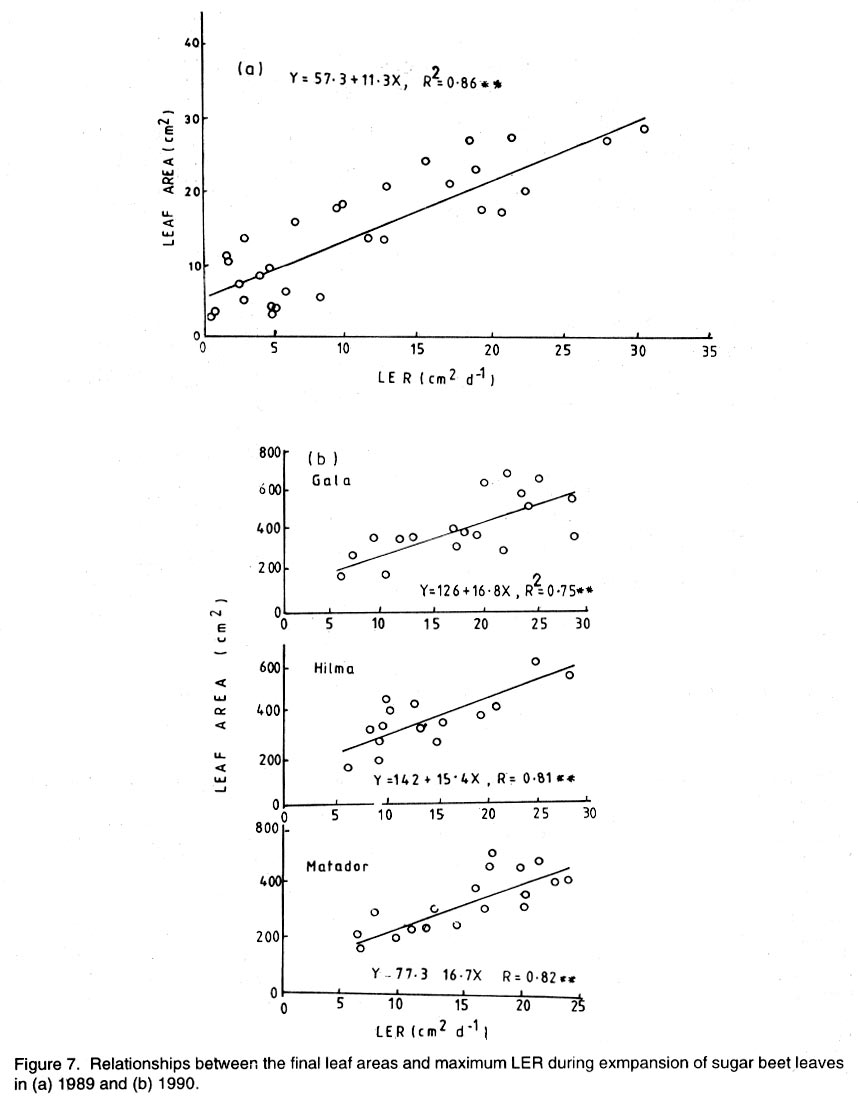
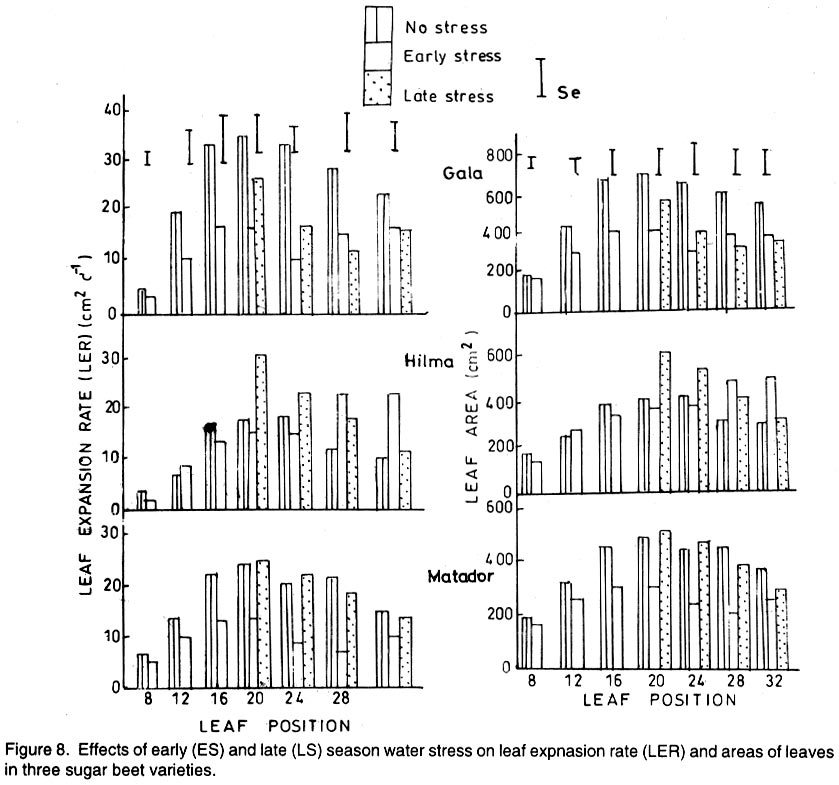
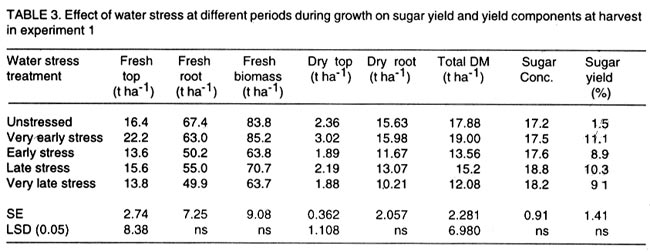
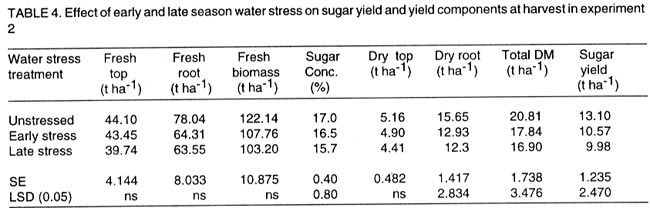
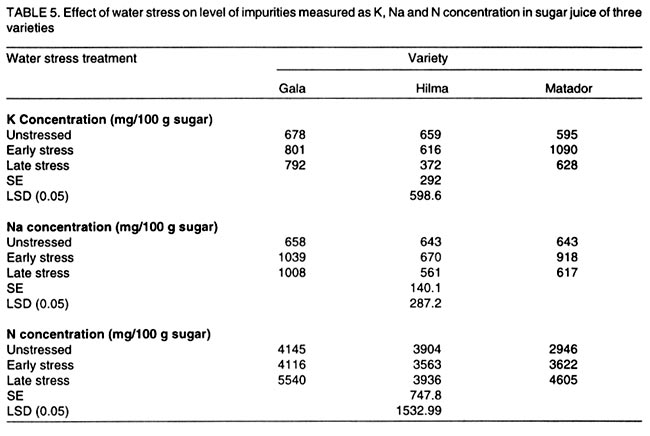
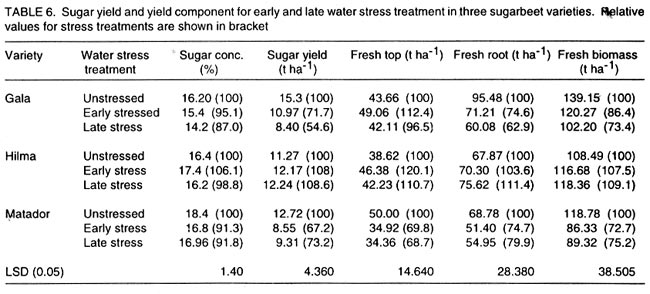
Kwara State, Nigeria (Received 18 October, 1999; accepted 27 November, 2001) Code Number: cs02006 ABSTRACT There are conflicting reports about the sensitivity of sugarbeet (Beta vulgaris L.) to water stress during different growth stages. Although, it is generally believed that the crop benefits from irrigation, opinions still differ as to which growth stage irrigation should be applied. This study evaluated the responses of sugarbeet leaf growth, sugar yield and yield components to soil water deficit imposed at various periods during growth in a glasshouse. Leaf growth showed high sensitivity to soil water deficit and responses varied with periods at which the deficit occurred. Water deficit early in the growing season had larger effects on leaf growth, leaf extension rate (LER), area of individual leaf, and leaf area index (LAI). Mid- or late-season soil water deficit showed relatively smaller effects on leaf growth. Re-watering resulted in compensatory leaf growth in early stressed plants. Both early (ES) and late (LS) soil water deficit decreased sugar yield and sugar concentrations. However, there were no significant (P<0.05) differences in the severity of effects of ES and LS, thus, no evidence to suggest that any particular growth stage is more sensitive to water stress than the other. Key Words: Beta vulgaris, irrigation, leaf area index, leaf extension rate, water stress RÉSUMÉ Il existe des rapports conflictuels concernant la sensibilité de la betterave à sucre (Beta vulgaris L.) aux stress d'eaux à différentes étapes de la croissance. Même s'il est généralement admis que les plantes profitent de l'irrigation, les opinions continuent à différer concernant le moment d'application. Cette étude a évalué l'accroissement de la feuille de la betterave à sucre, son rendement en sucre, et les autres composantes du rendement à la carence en eau imposée à des époques différentes de sa croissance dans en serre. La croissance des feuilles a montré une grande sénsibilité à la carence en eaux. La réponse a varié aussi avec la période à laquelle la carence s'est declarée. La déficience en eau précoce a produit un grand impact sur la croissance des feuilles, le taux d'accroissement, la surface foliaire individuelle, et l'indice de surface foliaire. A la mi ou vers la fin de la saison, la carence en eau dans le sol montra des moindres effets sur la croissance des feuilles. L'irrigation a eu un effet compensatoire sur la croissance des feuilles des plantes stressées précocement. Les deux déficits d'eaux, précoce et de fin de saison, ont diminué le rendement du sucre et la concentration en sucre. Cependant, la carence précoce et celle de fin de saison n'ont pas généré différents impacts apparent. Cependant, il n' y a pas assez d'evidences, pour affirmer qu'il existe d'étapes très sensibles que les autres à la carence en eau. Mots Cles: Beta vulgaris, irrigation, indice de surface foliaire, taux d'accroissement des feuilles, carence en eau Introduction There are conflicting reports about the sensitivity of sugarbeet to water stress conditions (Salter and Goode,1967; Brown, 1988; Davidoff and Hanks, 1989). Salter and Goode (1967) concluded that when sugarbeet is grown for root, there are no obvious indications that it is sensitive to moisture conditions at any particular stage of growth. However, the Food and Agricultural Organisation (FAO, 1977) later identified 3-4 weeks after emergence as a sensitive stage. Brown et al. (1987) suggested that this might apply more to dry climates, where plants depend wholly on irrigation. From available literature, there is evidence that sugarbeet benefits from supplementary irrigation even in places such as Britain which are less prone to drought conditions. Nevertheless, opinions still differ on which growth stage irrigation should be applied. Penman (1971) found that irrigation in June and early July enhanced early leaf growth but failed to increase sugar yield. A contradiction was, however, raised by Draycott and Massem (1977; 1978) that irrigation in June and July produced larger average sugar yield increases than in August and September. This was supported by other workers who showed that early drought (June and July) reduced yield more than late drought (August/September) (Brown et al., 1987; Brown, 1988; Dunham, 1988), thereby stressing the need for early irrigation. In a survey of severe drought damage of sugarbeet in some areas of Japan, Inoue et al. (1985) found that drought injury reduces root yield to 50% of a normal year, but did not affect sugar content. A similar report by Davidoff and Hanks (1989) showed that yields and relative yield exhibited a strong relationship with evapotranspiration (ET), but sucrose concentration was not significantly affected by irrigation. Simulation calculation with a dynamic model of yield formation in sugarbeet showed that root yield was enhanced by improved water supply, and that water stress at early stages of growth decreased yield more than stress at later stages because of retardation in leaf growth (Hoffmann, 1981). Kiss et al. (1985) have also shown that drought during the first half of the growing phase retards root formation, foliage and root development, thereby depressing sugar content. In sugarbeet, the final dry matter and sugar yields are known to be proportional to the total amount of radiation intercepted by crops during growth (Scott and Jaggard, 1978; Milford et al., 1980). Moreover, light interception is largely determined by leaf area index (Milford et al., 1985a). Drought stress has been shown to reduce the rate of leaf initiation (Clough and Milthorpe, 1975) and leaf expansion (Watts, 1974), thereby resulting in smaller leaf area which can adversely affect yield (Hsiao et al., 1976). Teresiak (1987) observed that drought caused wilting in unirrigated plots, decreased radiation absorption to about 62% and increased weed population. Similar reports from pot and plot trials showed that drought decreases the rate of leaf appearance and decreases the total assimilatory surface of leaves by up to 50% (Kazakov et al., 1988). In the present study, the sensitivity of growth and yields of sugarbeet varieties to soil water deficits applied at different periods during growth was evaluated. Materials and methods The study was conducted in an unheated glasshouse at the College Farm, Aber, University of Wales, Bangor, Gwynedd, LL57 2UW, UK, (53° 13'N, 4°9'W) during the summer periods of 1989 and 1990, in lysimeter tanks (100 cm x 100 cm) filled with sandy soil (pH 6.5). The sugarbeet varieties studied were Gala, Hilman and Matador, and the seeds, which were pelleted and treated as normal commercial seeds, were supplied by the British Sugar Company. The first experiment evaluated the effect of water stress at four growth stages on leaf growth and sugar yields of the variety Gala. In sugarbeet, as there are no distinct phenological growth stages as such (Milford, 1973), the choice of periods for stress application was largely arbitrary, the period between sowing and harvesting being divided into four equal parts and stress being applied by withholding watering for 14 days during each quarter, after which plants were irrigated to monitor recovery. Irrigation in unstressed plots was done every other day using garden hose fitted with roses. Thus, water stress was initiated in May (32 days after sowing (DAS)), July (79 DAS), August (110 DAS) and September (140 DAS), representing very early stress (VES), early stress (ES), late stress (LS), and very late stress (VLS), respectively. Seeds were sown in April, 1989, at a seed rate of 15 plant m-2. Each plot consisted of five rows spaced at 20 cm by 30 cm. Each plot received fertiliser at 100 kg N, 200 kg K and 50 kg P per hectare. Nitrogen was applied in two equal halves at planting and 12 weeks after sowing (WAS), with all K and P applied at planting. Weed control inside the tanks was achieved by hand rogueing at two weekly intervals, while weeds around the tanks as well as in the area surrounding the glasshouse were controlled by the application of a single dose of sodium chlorate made two weeks before sowing. During each water stress and recovery periods, the number of leaves appearing were recorded weekly and leaf expansion rate (LER) was measured in selected leaves. Estimates of LER were made based on two dimensional linear measurement using a ruler. Leaf length to the nearest millimeter was measured from the junction of lower lateral vines to the apex and leaf breath at its widest section. Leaf area was then calculated as the product of length x breath x a factor of 0.75 (Miford et al., 1985a), and leaf expansion rate (cm2d-1) was calculated as described by Tanguiling et al. (1987). Soil water content in both stressed and unstressed plots was measured every other day using a moisture meter (model 5910, soil moisture Equipment Corp, Santa Barbara, Calif. 93105, U.S.A.) connected to the terminals of gypsum electrical conductivity blocks placed at 30 cm below the soil surface. Mid-day leave water potential was measured on fully expanded, non-scenescent leaves, at the end of the stress period prior to rewatering and at 5 days after rewatering, using a pressure chamber located in the glasshouse. Sugar yield and yield parameters were recorded at harvest. Ten central plants in each plot were uprooted and the top was separated from the root at the level of the crown. Roots were washed to remove soil. The tops and roots were then weighed separately to obtain fresh weight. A 20% sample was taken from the thoroughly mixed top for oven drying at 100 oC to determine dry matter yield. Also, fresh root samples were taken for sugar determination using the procedure suggested by the Allscott sugar factory of the British Sugar Plc. Dried root samples were grounded and analysed for N using Kjeldah method (AOAC, 1955), K and Na by flame photometry (Isaac and Kerber, 1971). In the second experiment, the effect of water stress applied at two periods during growth was evaluated, on leaf growth and yield of three sugarbeet varieties. As in the first experiment, the growth stages for water stress application were arbitrarily chosen, the period between sowing and harvesting being divided into two equal halves. Water stress was then applied for 28 days during each period and, thereafter, plots were watered to monitor recovery. Water stress was initiated in June (68 DAS) and in August (120 DAS), representing early stress (ES) and late stress (LS), respectively. The evaluated varieties included Hilma and Matador, in addition to Gala used in experiment 1. Seeds were sown in April, 1990. Agronomic management and data collection were the same as described in the first experiment. The two experiments were designed as a randomised complete block with 4 and 5 replicates in experiments 1 and 2, respectively, and with treatments in a split-plot arrangement. Main-plot were three water stress treatments and subplots were sugarbeet varieties. All data were subjected to analysis of variance (ANOVA) with the appropriate models using GENSTAT Statistical software. Significantly different means compared using Least significant difference (LSD) P<0.05 probability level. Results and Discussion The weekly averages of daily maximum and minimum temperatures in the glasshouse during plant growth are presented in Figure 1. In 1989, temperature increased after sowing, reached peak values at 65 DAS, then declined steadily until harvest. In the early stress periods, plants experienced higher temperatures than the late stressed ones. However, in 1990, the maximum and minimum temperatures increased from 24.4 and 5.4 oC at 7 DAS, to peak values of 38.4 and 16.9 oC at harvest. These show that early stressed plants experienced slightly lower temperatures than the late stressed ones. However, with the eventual stress relief, plants recovering from ES experienced higher temperatures than those recovering from LS. Soil water content in irrigated treatments was maintained at near maximum water capacity (0.28 M3M-3) by daily irrigation using garden hose fitted with roses, while that in water-stressed treatment declined continuously from full capacity to about 0.08, 0.06, 0.15 and 0.18 M3M-3 and VES, ES, LS and VLS, respectively, prior to re-watering (Fig. 2a). Similarly, water content of soil in stressed plots declined continuously from about 0.32 M3M-3 to about 0.06 and 0.05 M3M-3 for ES and LS, respectively, before re-watering in experiment 2 (Fig. 2b). Mid-day leaf water potentials were lower in the water stressed plants with significant differences with VES, ES and LS, but not with VLS (Table 1), when soil water deficit decreased leaf water potentials by an average of -0.19, -0.58 and -0.63 Mpa prior to re-watering. During the VLS, leaf water potentials were lower for both water stress treatments. This was probably the result of low osmotic potentials. Re-watering of the water-stressed plants resulted in a gradual return of leaf water potentials to values similar to those of the unstressed control at all periods. The delay in full recovery of the leaf water potential may have been due to the time required for depletion of accumulated solutes that are associated with lower osmotic potentials under drought stress. Leaf growth responses to water stress. Leaf appearance rate and number of leaves appeared per plant were not significantly affected by water deficit at any period of growth (Table 2). This was in line with the results of Morton and Watson (1948) who showed that the rate of formation of new leaves was the same in sugarbeet plants exposed to severe and continuous stress in plots as in fully-watered plants. Leaf appearance rates were similar for all varieties evaluated in this study (Table 2). This is in agreement with previous findings by Milford and Riley (1980), who showed lack of significant varietal differences in the rate of leaf appearance in sugarbeet. However, leaf expansive growth showed high sensitivity to water stress, decreasing when water was limiting and resuming when water was more available (Figs. 3, 4 and 5). Other workers have shown that leaf growth in sugarbeet decreases with increasing water deficits (Miford et al., 1985b; McCree and Richardson, 1987). High sensitivity of leaf growth has also been reported for other species e.g., cotton (Gossypium hirsutum L.) (Rosenthatl et al., 1987; Davies and Xhang, 1991; Fernandez et al., 1996), sunflower (Helianthus annus L.) (Sandras et al., 1993), corn (Zea mays L.) (Acevedo et al., 1971), sorghum (Sorghum bicolor L.) (Fernandez, 1989) and cowpea (Vigna unguiculata L) Walp.) (McCree and Richardson, 1987). The expansion growth tends to be more sensitive to water stress than the meristematic process involved in leaf initiation, possibly because the short meristem and young leaves maintain higher water potential than other parts of the plant during periods of water stress (Lawlor and Milford, 1973). This is probably due to the fact that the apical activity of mature sugarbeet is less sensitive to water stress (Milford et al., 1985c), although water stress was known to restrict cell division in sugarbeet seedling leaves. Leaf growth results showed ontogenetic differences in response to water stress with the magnitude of the effects varying with leaf position. The effects of water stress on leaf expansion rate (LER) were greater on earlier (lower) leaves than on the later (upper) leaves (Fig. 6). Milford et al. (1985b) have shown similar differences in leaf response at different positions to water deficit. The ontogentic changes of leaf size in response to water stress observed in this study followed closely the seasonal changes in air temperature in the glasshouse (Fig. 1), which also affected soil moisture content (Fig. 2). For instance, LER was decreased by 79% by very early stress (VES) when mean temperature was 26.2oC and soil water content decreased to about 0.06 M3M-3 prior to re-watering, thereby resulting in strong positive correlations between temperature and maximum LER (r = 0.76, P<0.05), and the final leaf area (r = 0.86, P<0.01). Also, the data for both the stressed and unstressed plants in experiment 1 showed that the final leaf area was strongly associated with the maximum LER (Fig. 7a). These results are in agreement with those of earlier workers who have demonstrated that changes in leaf size were mostly associated with changes in the rates of expansion (Miford and Riely, 1980; Miford et al., 1985b) and that warmer air temperatures induce faster absolute rates of expansion (Milford et al., 1985a, c). Similarly, in experiment 2, the final leaf area was strongly associated with LER in the three varieties (Fig. 7b). Results also show that LER was consistently higher in Gala than in the other varieties at all leaf positions, hence, the variety always has larger leaves (Fig. 8). The data in Figure 8 show that, while both early and late water stresses had significant effects on Gala leaf growth decreasing from node 8 to node 32, only ES significantly decreased leaf growth in Matador. However, neither ES nor LS had a significant effect on leaf growth in Hilma. Similarly re-watering at the end of stress period had no significant effects on leaf growth recovery in Gala or Matador, but significantly improved growth in Hilma. These results indicate that leaf growth measurement could be very useful for distinguishing between genotypes in their responses to water stress. A frequently observed effect of recovery from water stress is a more rapid growth and development than in the unstressed controls, usually termed 'stored' or compensatory' growth (Kramer, 1983). Results of this study show that on relief of stress, LER recovered rapidly to values equal to or larger than those of the unstressed plants (Figs. 3 and 5). This is in line with the observation of Owen and Watson (1956), who reported that small amounts of rainfall after prolonged drought cause large increase in leaf growth of unirrigated sugar beet and these plants temporarily grow faster than the irrigated ones that had never been subjected to severe water stress. Similar observations were made by Miford et al. (1985b), who reported that after heavy rainfall, leaves resume expansion in the unirrigated crop and the extent of this growth was the same as in the irrigated crop. Bunce (1977) and Wenkert et al. (1978) found that reduction in growth rate caused by water deficit was made up by rapid 'compensatory' growth when leaves regained turgor. However, Hsiao and Acevedo (1974), explaining similar results in maize (Zea mays L.) leaf growth, had earlier suggested that the apparent faster growth after stress resulted from slowing of growth of the control leaves upon maturation, but definitely not from faster growth by stressed leaves than in the control leaves at the same development stage. In spite of the large increase in LER following re-watering, there were no appreciable changes in the size of leaves of previously stressed plants (Figs. 3, 4 and 5). These results corroborated with earlier observations that the most damaging feature of reduction in leaf area due to water stress is that the effect is permanent (Begg and Turner, 1976) although in the case of sugarbeet, there is scope of compensation via production of new leaves on relief of stress. This was clearly demonstrated in this study with the early stressed plant having similar LAI (5.13) as the unstressed control (5.52) at the final determination. Effect of water stress on yield and yield parameters. Table 3 shows that root and sugar yields were not significantly (P<0.05) decreased by water stress at any period in the first trial, although the magnitudes of the decrease were higher with late stress. It has been demonstrated that the ability of a plant to compensate for a reduction in leaf area allows the plant to recover from water stress with no loss of yield (Rawson and Turner, 1982). This was demonstrated by plants stressed very early (VES) in this study, which showed greatest effects of water stress on leaf growth, but lesser effects on root and sugar yields. The greater response of leaf growth was possibly a result of the prevailing higher temperature which aided leaf growth in unstressed plants. However, relief of VES resulted in compensation of leaf growth, which allowed the plant to recover fully. Moreover, the smaller effect of VES on root yield can also be explained by the fact that the initiation of secondary root thickening had not actually started by the time stress was applied. So the compensatory top growth after relief of stress actually helped the process of root thickening. Stress, however, occurred at the time of secondary root thickening with ES, LS and VLS treatment, hence, the greater effects on root yield. It should be pointed out that water stress was applied for 14 days during each period and so was of short duration. The result, therefore, demonstrated that sugarbeet can recover from water stress of short duration at any growth period, with little or no yield loss, thereby being in agreement with the observation that there is a possibility for full recovery under a short period of stress (Acevedo et al., 1971). A longer stress period would be required if any significant genotypic variations are to be obtained in screening for drought tolerance of sugarbeet. Consequently, stress was applied for 28 days in experiment 2. Results of the second trial showed that mean sugar yield reductions due to water stress across varieties were 19 and 24% for ES and LS, respectively (Table 4). Yield component analysis showed that clean root yield was decreased by 18 and 19% by the same treatment, respectively. These results suggest that the main factor for yield reductions was decrease in root yield, although reduced sugar concentration may have been an important factor under LS condition. Both ES and LS decreased sugar concentration at harvest, though, Brown et al. (1987) reported that late drought produced a higher sugar concentration, while the early one caused a lower percentage. The apparent disagreement of these results with those of Brown et al. (1987) may be due to re-watering of late stressed plants in this study, which possibly diluted the accumulated sugar during stress. Draycott and Messem (1978) showed that autumn rain reduces sugar percentage in crops subjected to increased water deficit, but does not normally affect crops which have been regularly irrigated. The concentrations of K, Na and amino N present as impurities in extracted root sap (Brown et al., 1987) have been shown to be inversely related to the amount of extractable sugar (Last et al., 1983). Results of the present study show that sugar concentration was negatively correlated with K (r = -0.37, P<0.01) and Na (r = 0.60, P<0.01) concentrations in the root. Although amino N was not determined in this study, total N was measured and showed a similar negative relationship with sugar concentration (r = 0.70, P<0.01). The concentration of melassigenic elements was not significantly different for both stressed and unstressed plants (Table 5), thereby suggesting higher levels of impurities was an important factor associated with sugar reductions due to water stress in this study. There were no apparent differences in the severity of effects of early and late season water stress in the present study. This contrasts with the result of Brown et al. (1987) and Penman (1971) that early drought is more detrimental to sugar yield. Nevertheless, our results are in agreement with earlier reports by Salter and Goode (1967) that when sugar beet is grown for its root, there are no obvious indications that it is especially sensitive to moisture conditions at any particular stages of growth. Varieties differed markedly in response to water stress during the periods (Table 6); ES and LS decreased yield by 28 and 45%, respectively, in Gala and by 33 and 27% in Matador. However, both ES and LS treatments increased sugar yield by 8 and 9% in Hilma. This is in conformity with our results of the leaf growth of these varieties (Fig. 8), and this highlights the suitability of leaf growth measurement as a drought resistance indicator in sugarbeet. The severity of the effects of LS was relatively greater on both root yield and sugar concentration, and, hence, on sugar yield of Gala than those of ES. In contrast, ES had relatively large effects on sugar yield and yield components of Matador. This differential responses of sugar beet varieties to water stress could, in part, offer some explanation for the conflicting reports on the severity of the effects of water stress at different growth periods, since the various workers had not used the same varieties. REFERENCES
©2002, African Crop Science Society The following images related to this document are available:Photo images[cs02006t6.jpg] [cs02006t5.jpg] [cs02006f3.jpg] [cs02006f8.jpg] [cs02006t4.jpg] [cs02006f5.jpg] [cs02006t2.jpg] [cs02006f2.jpg] [cs02006f1.jpg] [cs02006f6.jpg] [cs02006f7.jpg] [cs02006t3.jpg] [cs02006t1.jpg] [cs02006f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}