 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 10. No. 1, 2002, pp. 99-110 INTEGRATED MANAGEMENT OF GROUNDNUT ROSETTE DISEASE P. SUBRAHMANYAM, P.J.A. VANDER MERWE, A.J. CHIYEMBEKEZA and S. CHANDRA1 International Crops Research Institute for the Semi-Arid Tropics (ICRISAT),
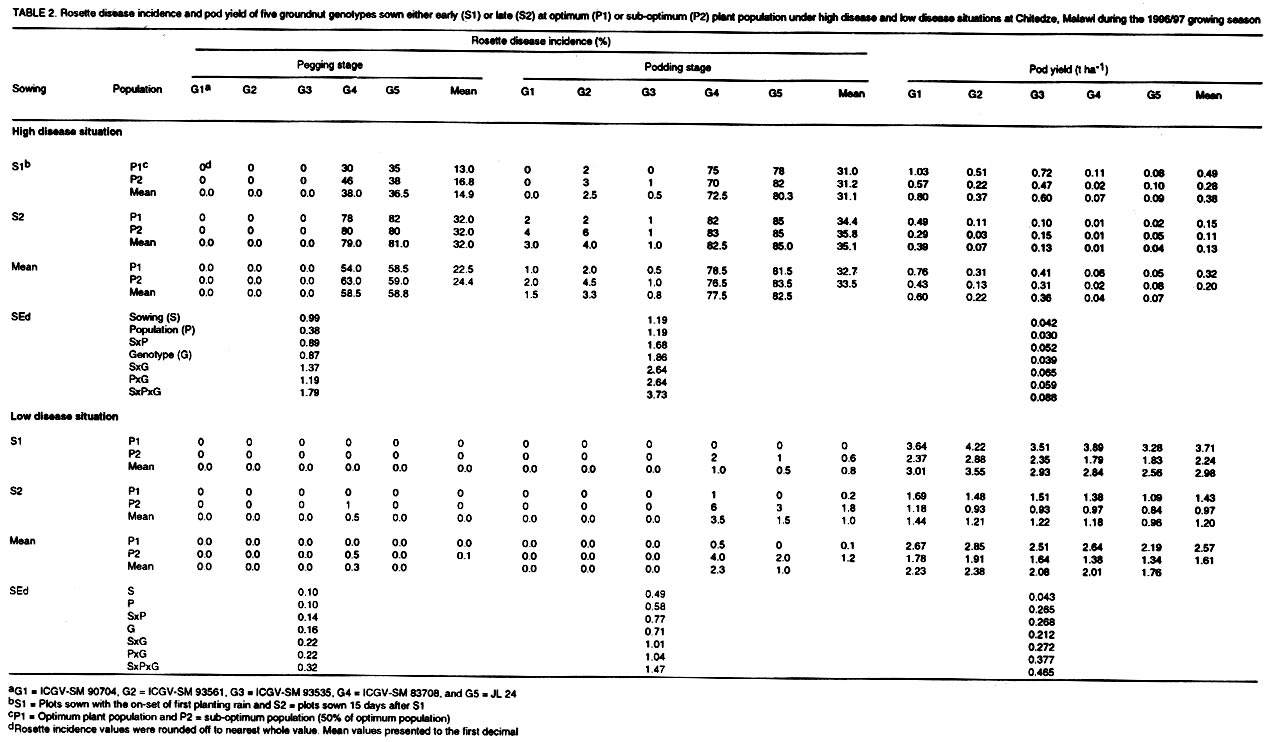
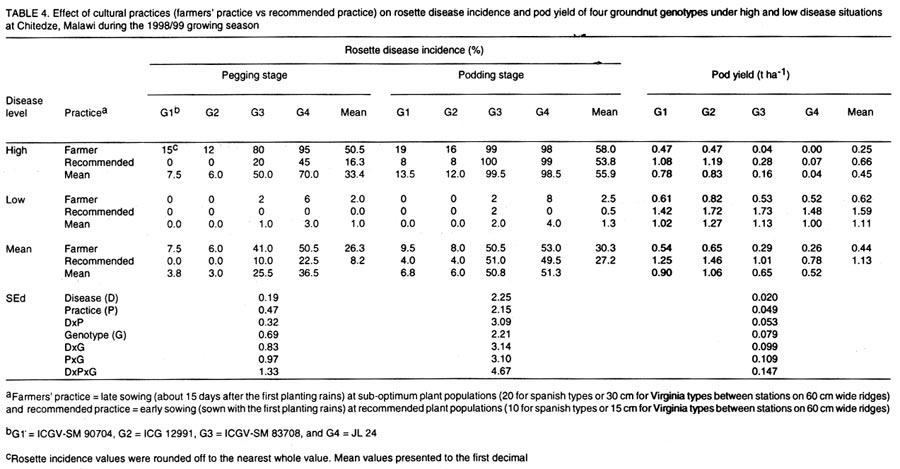
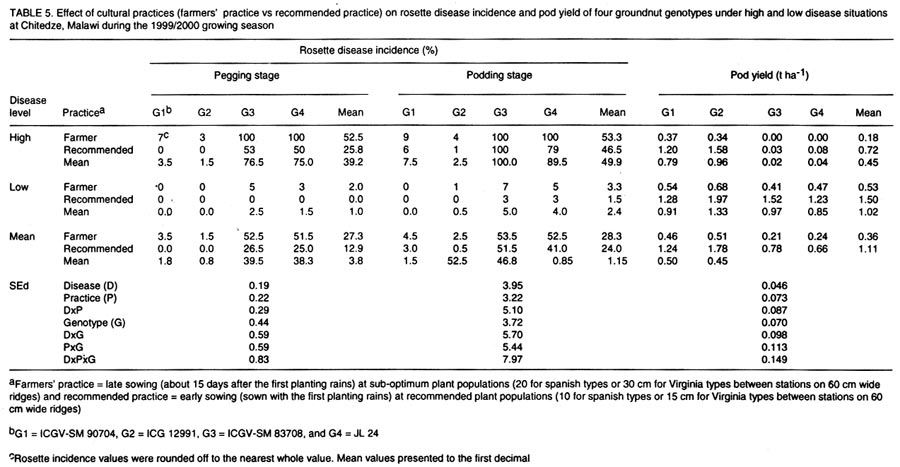
(Received 15 August, 2000; accepted 10 November, 2001) Code Number: cs02009 ABSTRACT An investigation was conducted to study the effects of cultural practices and host-plant resistance on rosette disease incidence and pod yields of elite groundnut genotypes during four growing seasons in Malawi. The effects of genotypes and cultural practices, and their interactions on rosette disease incidence were significant at pegging stage under high disease pressure in all seasons. The genotypic differences in disease incidence were spectacular under high disease infestation in all seasons. Resistant genotypes gave higher pod yields than susceptible genotypes under high disease incidence, irrespective of sowing time and plant population. However, pod yields were higher for early sown treatments at optimum plant population. There were no marked differences in pod yield between genotypes under low disease pressure. Overall, the combination of resistant genotypes with early sowing at optimum plant population exhibited the best performance under high disease infestation. Although the resistant genotypes gave the highest pod yields under high disease infestation, pod yields were consistently lower than the yields under low disease situation probably because of their susceptibility to groundnut rosette assistor virus (GRAV) component. Key Words: Arachis hypogaea, cultural practices, genotypes, host-plant resistance, optimum plant population RÉSUMÉ Une enquête visant à evaluer les effets des pratiques agricoles et la résistance des plantes - hôtes sur l'incidence de la rosette et le rendement en gousses des variétés - elites d'arachide, a été réalisée, durant 4 saisons culturales, au Malawi. Les effets de génotypes et de pratiques agricoles, et leurs interactions sur l'incidence de la rosette, ont été significatifs, durant la période de stabilisation, lorsque les plantes sont soumises sous une haute pression de la maladie. Les différences génotypiques sur l'incidence de la maladie, ont été spectaculaire, durant toutes les saisons lorsque les plantes sont soumises sous une haute pression de la maladie. Les génotypes résistants ont donné des rendements en gousses plus élévés par rapport aux susceptibles, quelque soit la période de semis et la densité des plantes. Cependant, les rendements étaient élévés pour un semis précoce réalisée à la densité des plantation optimale. Au point de vue de rendement en gousses, des différences non significatives, entre les génotypes, étaient obsrvées lorsque les plantes étaient soumises sous faible pression de la maladie. Les meilleures performances de rendement, lorsque les plantes sont soumises sous hautes pressions, ont été observées, lorsque l'on combine génotypes résistants et semis precoces. Malgré que les génotypes résistants ait donné, un rendement élévé en infection élévée, les rendements étaient moins élévés par comparaison à la situation de faible pression, probablement à cause de leur sensibilité aux composantes du GRAV. Mots Clés: Arachis hypogaea, résistante des plantes- hôtes, pratiques agricoles INTRODUCTION Rosette is the most destructive viral disease of groundnut (Arachis hypogaea L.) in Africa. The disease is endemic to sub-Saharan Africa and its off-shore islands (Reddy, 1991; Subrahmanyam et al., 1991, 1997; Naidu et al., 1999). Two main forms of the disease, chlorotic rosette and green rosette have been described based on symptoms (Gibbons, 1977). The disease is transmitted by the aphid, Aphis craccivora Koch (Homoptera: Aphididae)(Okusanya and Watson, 1966), and is caused by a complex of three agents: groundnut rosette virus (GRV), genus Umbravirus (Murant et al., 1995) and its satellite RNA (sat. RNA)(Blok et al., 1994) and groundnut rosette assistor virus (GRAV), genus Luteovirus (Casper et al., 1983; Reddy et al., 1985; Murant, 1989). Rosette disease has been and continues to be responsible for serious losses to groundnut production in Africa. For example, the rosette epidemic in 1994/95 in central Malawi and eastern Zambia destroyed the crop to such an extent that the total area of groundnut grown in Malawi fell from 92,000 ha in 1994/95 to 65,000 ha in 1995/96. Losses in Zambia were estimated at US$ 5 million that year (Subrahmanyam et al., 1997). Overall yield loss due to rosette disease in Africa was estimated at about US$ 156 million per annum. The potential yield gain due to management of the disease was estimated at about US$ 121 million (ICRISAT, 1992). Management of groundnut rosette by insecticidal control of the vectors has been known since the mid-1960s (Davies, 1975a, b). However, chemical control is not economically feasible to smallholder farmers in Africa, mainly because of resource limitation. Chemical control requires careful monitoring of the vectors and may not be feasible at farmer level. With increased concern for environmental preservation, this practice now has further disadvantages. Information is also available on the control of groundnut rosette using cultural practices, particularly manipulation of sowing date and plant density. Several researchers have demonstrated that groundnut rosette can be reduced when the groundnut crop is sown early in the season and at optimum plant densities (Subrahmanyam et al., 1992; Subrahmanyam and Hildebrand, 1994; Naidu et al., 1999). Intercropping groundnuts with beans is reported to decrease rosette incidence in Malawi and Uganda, and similar effects were found when groundnuts were intercropped with maize in the present Central African Republic (Subrahmanyam et al., 1992). Pioneering research on development of groundnut varieties with resistance to rosette was carried out in West Africa. Sources of resistance to rosette were first discovered in Senegal in 1952 in germplasm originating from the frontier region between Burkina Faso and Cote d'Ivoire (Catherinet et al., 1955). These sources formed the basis for rosette resistance breeding programmes throughout Africa and have contributed to the development of several high-yielding, rosette-resistant groundnut varieties in Senegal, Burkina Faso, Nigeria, Malawi, Mozambique, and Uganda and to a lesser extent in other countries in Africa. However, most of the rosette-resistant varieties are late maturing and are not suitable to some production systems in Africa where the rainy season is short. In recent years, a large number of high-yielding, agronomically acceptable, medium- to long-duration Virginia type (e.g., ICGV-SM 90704) and short-duration Spanish type (e.g., ICG 12991) groundnut varieties with resistance to rosette disease have been developed (Chiyembekeza et al., 1997; vander Merwe and Subrahmanyam, 1997; Reddy and Subrah-manyam, 1997; vander Merwe et al., 1999; Naidu et al., 1999). In this paper, the effects of sowing time (early and late) and plant population (optimum and sub-optimum) on rosette disease incidence and pod yields of some elite high-yielding groundnut genotypes (rosette-resistant and rosette susceptible) under two disease situations (high and low) during the 1996/97 and 1997/98 growing seasons at Chitedze, Malawi are reported. A combination of cultural practices, early sowing at optimum population ('recommended practice') and late sowing at sub-optimum population ('farmers' practice') on rosette incidence and on pod yields was also investigated in1998/99 and 1999/2000 and the results are presented in this paper. MATERIALS AND METHODS Field trials were conducted during the 1996/97, 1997/98, 1998/99, and 1999/2000 growing seasons (December-April) at Chitedze Agricultural Research Station located 16 Km west of Lilongwe, Malawi at 140S and 33045'E with an altitude of 1149 m above sea level. The genotypes used in the experiments are described in Table 1. Seeds were treated with the fungicide thiram at 3 g kg-1 seed before sowing. All trials were conducted under rainfed conditions and were fertilised with 40 kg P2O5 ha-1 as basal application of single super phosphate. First and second growing seasons (1996/97 and 1997/98). The effects of sowing dates (early and late) and plant populations (optimum and sub-optimum) on rosette disease incidence and pod yields were investigated on five groundnut genotypes (three rosette-resistant and two rosette-susceptible) in 1996/97 and on seven groundnut genotypes (five rosette-resistant and two rosette-susceptible) in 1997/98. Each genotype was sown at the on-set of planting rains (early sowing) or 15 days after the first sowing (late sowing) at a recommended spacing of 10 cm for Spanish types or 15 cm for Virginia types (optimum population) or 20 cm for Spanish types and 30 cm for Virginia types (sub-optimum population) along 60 cm wide raised ridges. In both seasons, the trials were conducted under two disease infestation levels (high and low) by artificial inoculation of infector rows (Bock and Nigam, 1988; Subrahmanyam et al., 1998). A chlorotic rosette culture maintained in the glasshouse was used to create high disease pressure in the field. Infector rows of a rosette susceptible variety (cv. Malimba) were arranged throughout the trial, two infector rows flanking each plot. Potted spreader plants (cv. Malimba) showing severe rosette symptoms and heavily infested with aphids were raised in the glasshouse and transplanted in the infector rows (2 per 6 m row) 10 days after sowing. High disease pressure was achieved by further infesting the viruliferous aphids raised in the glasshouse until all plants in the infector row were infested with aphids. To achieve low disease situation, only non-viruliferous aphids were used to infest the infector row. Plots were arranged in a three-replicate split-split-plot design with dates of sowing as main-plots and levels of plant population as sub-plots and genotypes as sub-sub-plots (14.4 m2). Third and fourth growing seasons (1998/99 and 1999/2000). The effects of recommended cultural practices (early sowing at optimum population) and farmers' practice (late sowing at sub-optimum populations) on rosette disease incidence and pod yields of four genotypes (two rosette-resistant and two rosette-susceptible) were investigated in 1998/99 and 1999/2000, as described above. Each genotype was sown at the on-set of rains at a recommended spacing of 10 cm for Spanish types or 15 cm for Virginia types (recommended practice) or 15 days after the first sowing at a sparse spacing of 20 cm for Spanish types or 30 cm for Virginia types (farmer's practice) along 60 cm wide raised ridges. In both seasons, the trials were conducted under two disease infestation levels (high and low), as described above. Plots were arranged in a three-replicate split-split-plot design with disease levels as main plots, cultural practices as sub-plots and genotypes as sub-sub-plots (27.0 m2). In all seasons, each plot was assessed for rosette disease incidence initially at pegging stage and finally at pod filling. The total number of plants in each plot and number of plants showing chlorosis with severe stunting were counted and the percentage of disease incidence was computed. Plots were harvested at optimum maturity by hand and yields of dried pods were recorded. The data on rosette disease incidence and pod yield were analysed using a fixed-effects analysis of variance (ANOVA) model as per split-split-plot design. The data on pod yields reasonably satisfied the ANOVA assumptions of variance homogeneity, additivity, and normality. The rosette disease incidence data, being binomially distributed, were transformed to angular scale and both transformed and raw data subjected to ANOVA (Steel et al., 1997). However, the analysis revealed no differences in variations between the angular transformed values and the original data sets. Therefore, results for disease incidence are reported on the original scale. GENSTAT release 4.1 statistical computing package was used to conduct the analysis (Lane and Payne, 1996). RESULTS AND DISCUSSION Groundnut genotypes.The groundnut genotypes used in this study are described in Table 1. ICGV-SMs 90704, 93561, and 93535 are high-yielding, rosette-resistant breeding lines developed at ICRISAT, Malawi. Breeding line, ICGV-SM 90704 was released in Uganda (as Serenut 2) and Malawi. It is at the pre-release evaluation stage in Zambia and Mozambique. Lines ICGs 12988 and 12991 are high-yielding, rosette-resistant land races from India. Also, ICG 12991 is at pre-release evaluation in Malawi and Mozambique. Another line, ICGV-SM 83708 is a high-yielding genotype but susceptible to rosette. It was released in Malawi as CG 7, in Zambia as MGV 4, and in Uganda as Serenut 1. Lastly an introduced line, JL 24 is from India and has been released in Zambia (as Luena) and Malawi. Rosette disease incidence. High incidence of rosette was achieved by inoculating the infector rows with viruliferous aphids in all growing seasons, obtained from susceptible genotypes under 'high disease' situation. Infected plants were chlorotic and severely stunted, resulting in drastic reduction in pod yields. However, the disease incidence was very low in plots where infector rows were inoculated with non-viruliferous aphids (Tables 2, 3, 4, and 5). These differential disease situations (high and low) were ideal for studying the effects of cultural practices, genotypes and their interaction on rosette disease incidence and yields. It also provided an opportunity to examine the performance of rosette-resistant genotypes in the absence of the disease. This is especially important for a disease like rosette, which does not occur in epidemic proportions every year. Rosette epidemics occur only sporadically in most groundnut producing countries in Africa (Subrahmanyam et al., 1991, 1997). The 'high disease' infestation level in this study is comparable to severe epidemic situation. Although there was some disease development under the low disease situation, the incidence was very low and comparable to non-epidemic situation. Genotype (G) differences for rosette disease incidence were consistently highly significant (P <0.001) under high disease situation in both seasons of 1996/97 and 1997/98 and also under low disease situation for the 1997/98 season. The differences in sowing time (S) and plant population (P) were significant (P <0.01) at pegging stage under the high disease situation in both seasons and under low disease situation in 1997/98. The interaction effects of S x P, S x G, P x G, and S x P x G were highly significant (P <0.001) only at pegging stage under high disease situation in both seasons. These interaction effects were not significant under low disease situation. The differences in rosette incidence between resistant and susceptible genotypes under high disease situation were very spectacular in all seasons (Tables 2 and 3). All resistant genotypes consistently showed low disease incidence in both growing seasons under the high disease situation. The resistant genotypes consistently showed low disease incidence in both seasons irrespective of sowing time and plant population. Rosette incidence on susceptible genotypes at pegging stage was significantly lower in early sowing than in late sowing in both seasons. Effects of plant population on disease incidence were not significant at pegging except in late sowing in 1998/99. There were no significant effects of sowing time and population on disease incidence at the pod filling stage (Tables 2 and 3). The effects of disease level (DL), cultural practice (CP), genotypes (G), and their interactions DL x CP, DL x G, CP x G, and DL x CP x G on disease incidence were highly significant (P <0.001) at pegging for both seasons of 1998/99 and 1999/2000. At podding stage, only DL, G, DL x G were significant. As in previous seasons, the genotypic differences in disease incidence were very marked under the high disease situation in both seasons. Resistant genotypes showed much lower incidence of the disease than the susceptible genotypes. There were highly significant (P <0.001) disease incidence differences between farmers' practice and recommended practice under high disease situation at pegging stage but not at the pod filling stage (Tables 4 and 5). Pod yield. The effects of genotype on pod yields were highly significant (P <0.001) under high disease situation in both seasons of 1996/97 and 1997/98. However, under the low disease situation, it was significant (P <0.01) only in the 1999/2000 season. The effects of sowing date were not significant under the high but significant (P <0.01) under low disease pressure. The effects of plant populations were significant under both disease situations and seasons. The interactions of S x P, S x G, P x G, and S x P x G were not significant under low disease pressure in both seasons. However, under high disease pressure, the interactions of S x G and P x G in 1996/97 and S x P x G in 1997/98 were significant (P<0.05). Pod yields were substantially lower under high than under low disease pressure in both seasons. In general, the resistant genotypes gave higher pod yields than the susceptible ones under high disease pressure, irrespective of sowing time and population. However, pod yields were higher in early sowing than in late sowing situations in both seasons and disease situations. Yields were generally higher under optimum population than under sub-optimum population (Tables 2 and 3). Under high disease situations in 1996/97, the rosette-resistant genotype ICGV-SM 90704 gave the highest pod yield followed by ICGV-SM 93535. Under low disease pressure, there were no significant differences between genotypes in early sowing at the optimum population. The resistant genotypes gave high yields under early sowing at sub-optimum population and at late sowing at optimum population. In 1997/98, the rosette-resistant genotypes ICGs 12991 and 12988 gave higher pod yield in all treatments followed by ICGV-SM 90704 under high disease pressure. However, there were no significant genotypic differences in pod yield under low disease situation. The effects of DL, CP, G, and DL x G on pod yields were highly significant (P <0.001) in both seasons of 1998/99 and 1999/2000. The interaction effects of DL x CP in 1998/99 and CP x G and DL x CP x G in 1999/2000 were also significant. As for previous seasons, the yields were highest under low than under high disease pressure, irrespective of cultural practices in both seasons. Recommended practice gave higher pod yields than farmer practice under both disease situations and seasons. The resistant genotypes gave highest pod yields than the susceptible ones under high disease situation for both seasons (Tables 4 and 5). The susceptible genotypes produced very few or no pods under the high disease situation. There were no significant differences between genotypes under low disease situation in both seasons. However, ICG 12991 showed superior performance in 1999/2000 under recommended practice (Table 5). Several workers have reported that rosette incidence is generally low when groundnuts are planted early in the season using recommended seed rates and spacing to achieve optimum plant populations (Subrahmanyam and Hildebrand, 1994; Naidu et al., 1999). Cultivation of rosette-resistant genotypes is another best way of containing the disease. In the present study, all resistant genotypes consistently showed low disease incidence irrespective of sowing time and plant population. However, pod yields were higher at early sowing at optimum plant populations or when the crop was grown using recommended cultural practices both under high and low disease situations. Rosette incidence in susceptible genotypes was significantly lower at pegging stage in early sowing than in late sowing. Although, there were no significant effects of sowing time on disease incidence at pod filling stage, pod yields were generally higher in early sowing than late sowing situations. Plants in late sown crops were severely stunted. In general, early sowing at optimum population gave the highest pod yields in both seasons and disease situations irrespective of genotype. Overall, the combination of resistant genotype with early sowing at optimum population exhibited the best performance under high disease situation. Under the low disease situation, yields were highest for early sowing at the optimum population or when crop was grown using recommended practices and lowest under late sowing at sub-optimum population or when the crop was grown using farmers' practice in all seasons. The higher pod yield observed from early sowing at optimum plant populations or under recommended cultural practices can not be solely attributed to low rosette incidence. The higher yields obtained from these genotypes when grown under low disease situation, is a clear indication that the superior performance of these genotypes in early sowing at optimum population is due to a combination of climatic and biotic factors which play an important role in determining the yield performance of genotypes. It is interesting to note that although the resistant genotypes gave the highest yields under high disease situations, the yields were consistently lower than yields under low disease situations. The resistant genotypes used in this study are susceptible to groundnut rosette assistor virus (GRAV) (Subrahmanyam et al., unpubli.). Resistance to rosette in these genotypes is not absolute since a small proportion of plants or a few branches of plants in resistant genotypes showed rosette symptoms. Resistance to rosette in these genotypes appears to be against GRV which results in indirect resistance to its sat RNA, and thus, such genotypes do not develop symptoms. Although a high percentage of plants of these genotypes did not show visible symptoms in the field, they were considerably stunted under the high disease situation probably because of their susceptibility to GRAV. Although GRAV alone causes no visible symptoms, it appears to interact with other two agents in disease development (Naidu et al., 1998). The high yield reduction in resistant genotypes when grown under the high disease situation seems to be due to a combination of these factors. Hence, there is a need to reinforce resistance in these genotypes by incorporating resistance to GRAV and possibly to the vector to minimise the yield losses due to the disease. REFERENCES
©2002, African Crop Science Society The following images related to this document are available:Photo images[cs02009t5.jpg] [cs02009t2.jpg] [cs02009t4.jpg] [cs02009t3.jpg] [cs02009t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}