 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 10. No. 2, 2002, pp. 133-137 Inheritance of genes for leaflet shape and leaflet shape modifier in cowpea O.B. Oluwatosin Genetics and Molecular Biology Research Laboratory, Department
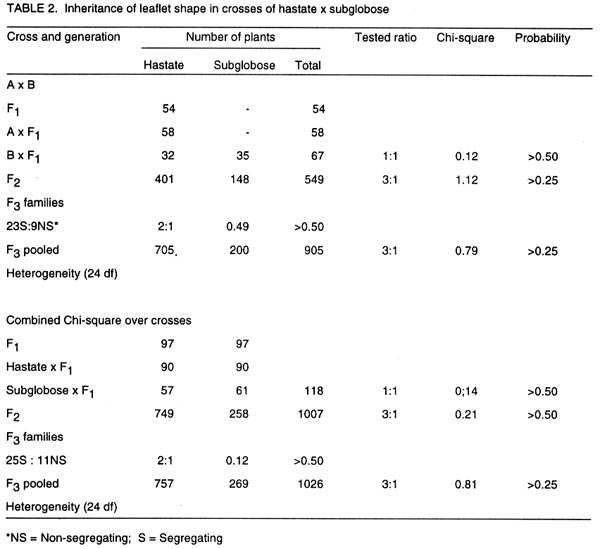
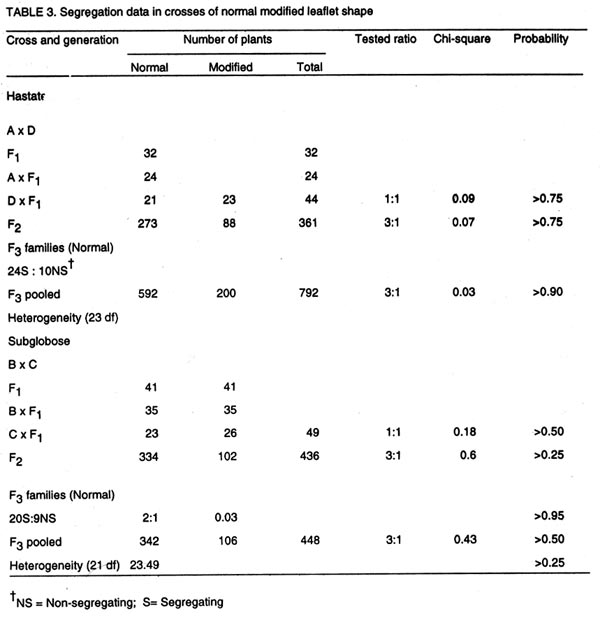
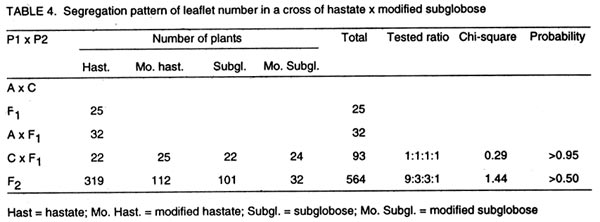
of Botany and Microbiology (Received 11 September, 1999; accepted 10 November, 2001) Code Number: cs02012 Abstract Crosses between five cowpea lines were used to determine the inheritance of genes for hastate and subglobose leaflet shapes obtained in the F2. Analysis of crosses between hastate x subglose leaflet shapes revealed that a single gene differentiated the two parental leaflet shapes. Outputs from the analysis of a modifier that was capable of modifying the hastate and subglobose leaflets shapes observed in the segregating population of a cross between IBS 876 x IBS 2625 indicated that the modifier was genetic and inherited in a recessive fashion. In crosses of hastate x modified hastate and subglobose x modified subglobose, F1 plants produced normal leaflet shapes while a 3 normal 1 modified leaflet shape ratio was obtained for the F2 generation. These results suggest that a recessive gene conditioned the leaflet shape modifier gene. Key Words: Hastate, modifier gene, Nigeria, subglobose, Vigna unguiculata Résumé Le croisement entre cinq génes du niebé étaient utilisés pour déterminer l'héritage des génes pour hastate et subglobose des formes des feuillets obtainues dans le F2. Les analyses de croisements entre hastate X subglobose sur les formes des feuillets ont revelé qu'un simple géne a différentié les deux formes des feuillets parentales. Les résultats des analyses d'un modifiant capable de transformer les hastates et les subgloboses des formes des feuillets observées dans la ségrégation des populations d'un croisement entre IBS 876 x IBS 2625 ont indiqué que le modifiant était génétique et hérité de façon récessive. Dans les croisements de hastate x hastate modifié et subglobose x subglobose modifié, F1 plantes ont produit des feuillets de formes normales alors que 3 normale et 1 modifié était observées pour la génération F2. Ces résultats suggérent qu' un géne récessif a contraint le modifiant génétique des formes de feuillets. Mots Clés: Hastate, modifiant génétique, Nigéria, subglobose, Vigna unguiculata INTRODUCTION Cowpea (Vigna unguiculata [L.] Walp) is grown primarily for its edible seeds in Africa (Wien and Summerfield, 1984), where it serves as a major source of essential nutrients (Oluwatosin, 1997a; 1998). However, cowpea leaves are also among the top three or four leaf vegetables in terms of preference in many parts of Africa, Asia and the Pacific (Barret, 1990; Oluwatosin, 1997b). About six different genes have been reported to condition leaf shape or size in cowpea (Krishnaswamy et al., 1945; Jindla and Singh, 1970; Kolhe, 1970; Ojomo, 1977; Fery, 1980). However, the inheritance pattern of hastate leaflet shape is largely unknown, with available literature indicating conflicting views. For example, Jindla and Singh (1970) proposed that four genes condition hastate leaflet shape, while Ojomo (1977) proposed a single gene. In addition, information on the relationship between the genes for hastate and subglobose leaflet shape is unavailable. Furthermore, a modifier, which was able to modify the hastate and subglobose leaflet shapes, was also observed in a segregating population of crosses involving lines with normal leaflet shapes. In view of the potential usefulness of the leaflet shape modifier as a morphological marker, knowledge of its inheritance pattern is necessary. This study reports on the inheritance of the hastate and subglobose leaflet shapes and a leaflet shape modifier. MATERIALS AND METHODS The names, origin and leaflet shapes of the cowpea lines used in this study are shown in Table 1. Reciprocal crosses were made between the cowpea leaflet shapes in screenhouse in May of 1993 at the University of Ibadan in Nigeria. Reciprocal F1 crosses were advanced to the F2 generation in the screenhouse and field in September of 1993. Backcrosses of F1 plants to the parents were also made in the screenhouse in December of 1993. Field evaluations were conducted at the University of Ibadan Teaching and Research Farm in the early and late seasons of 1994, and at the Department of Botany and Microbiology Nursery in the early and late seasons of 1995, 1996 and 1997. The parents, backcross, reciprocal F1 and F2 seeds were evaluated in 1994, 1995 and 1996, while progenies of selected lines were advanced to F3 generation in 1997. All field evaluations were conducted using a completely randomised block design with six replicates for each cross in the experiment. Each replicate consisted of a row of each of the parents, F1 and reciprocal F1, two rows each of the backcrosses to both parents and twelve rows of the F2. Forty F2 plants were randomly selected and advanced to F3 generation for each of the leaflet shapes. Only F3rows, which consisted of at least thirty-six plants, were scored. Field spacing was 60 cm between and 25 cm within rows. Diseases, weeds, insects and vertebrate pests were controlled using the appropriate cultural practices. Leaflet shapes were classified as hastate and subglobose and were scored at six weeks after planting by comparison with standard descriptors. When the hastate and subglobose leaflets were found to have deep inverginations at the apex, they were classified as modified hastate and modified subglobose leaflets shapes, respectively. The data obtained for the various generations were tasted for their goodness to the proposed genetic ratios using the chi-square test (Gomez and Gomez, 1984). RESULTS In crosses of hastate x subglobose (Table 2), F1 plants and progeny of backcrosses to the hastate parent produced hastate leaflets shape, indicating the dominance of hastate to subglobose leaflet shape. Backcross to the subglobose shape parents segregated in the ratio 1 hastate: 1 subglobose leaflet shape, while the F2 plants gave a 3 hastate: 1 subglobose leaflet shape ratio. These results indicate that a single dominant gene differentiated the hastate and subglobose leaflet shape in parents under study. The F2 individuals with subglobose leaflet shape bred true to type in the F3 and subsequent generation, while the segregation pattern of F2 plants with hastate leaflet shape showed a good fit to the 2 segregating: 1 non-segregating family ratio in the F3 indicates that hastate leaflet shape was monogenically dominant to subglobose shape. Hastate x modified hastate and subglobose x modified subglobose. Table 3 shows the segregation data of leaflet shapes in crosses of leaflet shapes x their respective modified forms. The F1 plants and progenies of the backcrosses to the hastate and subglobose leaflet shape parents had normal shapes and the F2 generation segregated for a good fit to the 1:1 and 3:1 normal to modified leaflet shapes ratio, respectively. This indicates that a single gene differentiated the normal leaflet shapes and their respective modified forms. Third generation (F3) families raised from F2 plants with the modified leaflets shapes bred true to type while 20 of the 29 families raised from F2 individuals with normal leaflet shapes segregated for a good fit to the 3 normal: 1 modified leaflet shape ratio. Hastate x modified subglobose. The F1 plants and progenies of backcross to the hastate parent in the cross of hastate x modified subglobose leaflet shapes, had hastate leaflet shape (Table 4) suggesting further that leaflet shape modification is controlled by a recessive gene. In the backcross to the modified subglobose leaflet shape parent, a 1 hastate: 1 modified hastate: 1 subglobose: 1 modified subglobose leaflet shapes was obtained. Moreover, F2 plants satisfactorily fitted the 9 hastate: 3 modified hastate: 3 subglobose: 1 modified subglobose leaflet shapes. These results indicate an independent segregation of the leaflet shapes and modifier genes. DISCUSSION This study revealed that hastate leaflet shape was inherited in a dominant fashion in agreement with earlier reports (Jindla and Singh, 1970; Ojomo, 1977). Results of the study also suggest that a single dominant gene conditioned the hastate leaflet shape and a single gene appeared to separate the hastate and subglobose leaflet shapes. The cross between the normal leaflet shapes and their modified forms indicates that the leaflet modifier is heritable and is monogenically recessive to the normal leaflets shape gene. The ratio of 9 hastate: 3 modified hastate; 3 subglobose: 1 modified subglobose leaflet shapes obtained in the cross hastate x modified subglobose confirms the independent assortment of the leaf shape and the leaflet shape modifier gene loci. This also indicated that two dominant independently assorting genes differentiated the two parents used in the cross. Ojomo (1977) proposed the gene symbol Ha for the hastate leaf shape gene. The symbol 1 m is proposed for the leaflet shape modifier gene reported in this study. The dominant allele Lm gives the normal leaflet form. The recessive allele ha gives the phenotype sub-globose. In conclusion, a single gene that was recessive to the hastate controls the subglobose leaflet shape. The leaflet shape modifier is also genetic and is inherited in a recessive fashion. Acknowledgement
The author is grateful to Dr. I. Fawole for the seeds of IBS 2625 and IBS 876 and to Dr. N.A. Amusa for editing the earlier manuscript. The author is also grateful to Fayemi, S.O. and Ajibade, K.O. of the Department of Botany and Microbiology, University of Ibadan, Ibadan, Nigeria for assisting with data collection. REFERENCES
©2002, African Crop Science Society The following images related to this document are available:Photo images[cs02012t2.jpg] [cs02012t4.jpg] [cs02012t3.jpg] [cs02012t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}