 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 10. No. 2, 2002, pp. 145-156 SELECTION OF NON-PATHOGENIC ETHYLENE-PRODUCING RHIZOBACTERIA FOR ACCELERATED DEPLETION OF Striga hermonthica SEED BANK M.O. AHONSI, D.K. BERNER1, A.M. EMECHEBE2, N. SANGINGA, and S.T. O. LAGOKE3 International Institute of Tropical Agriculture (IITA), PMB 5320,
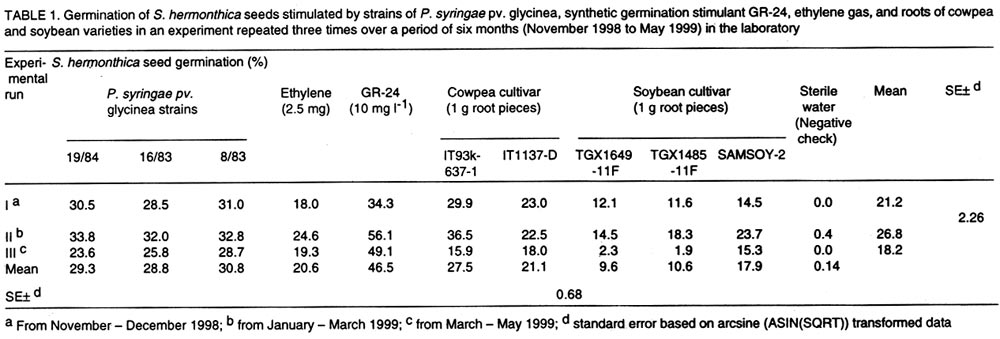
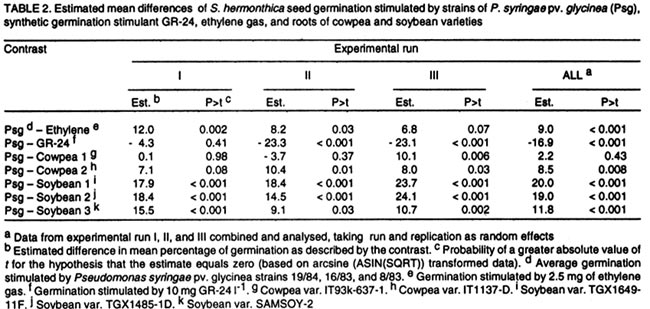
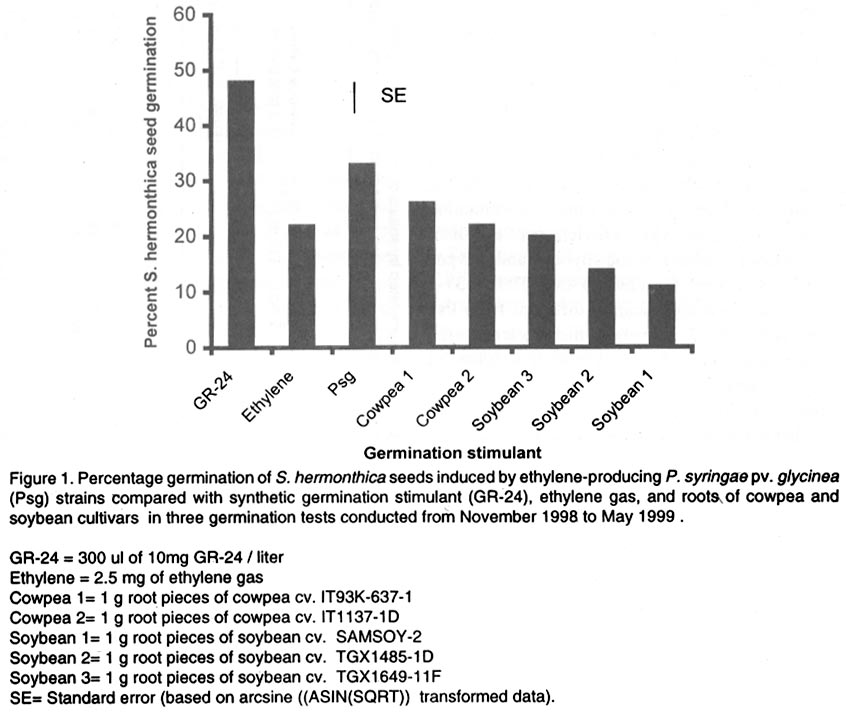
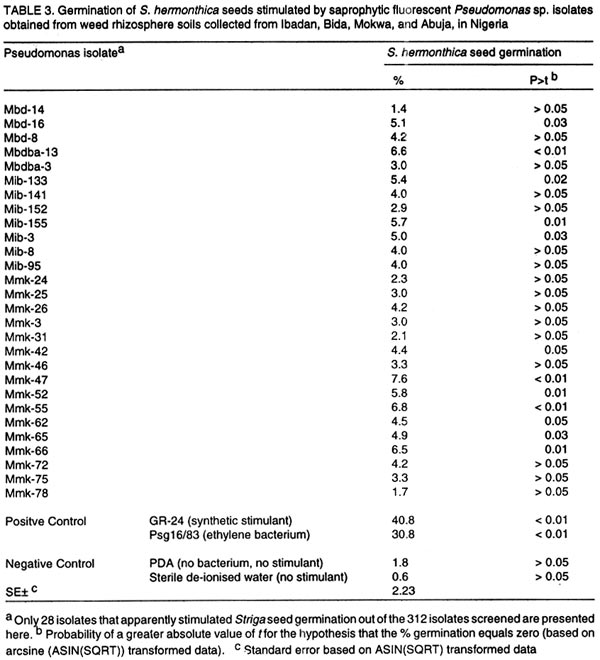
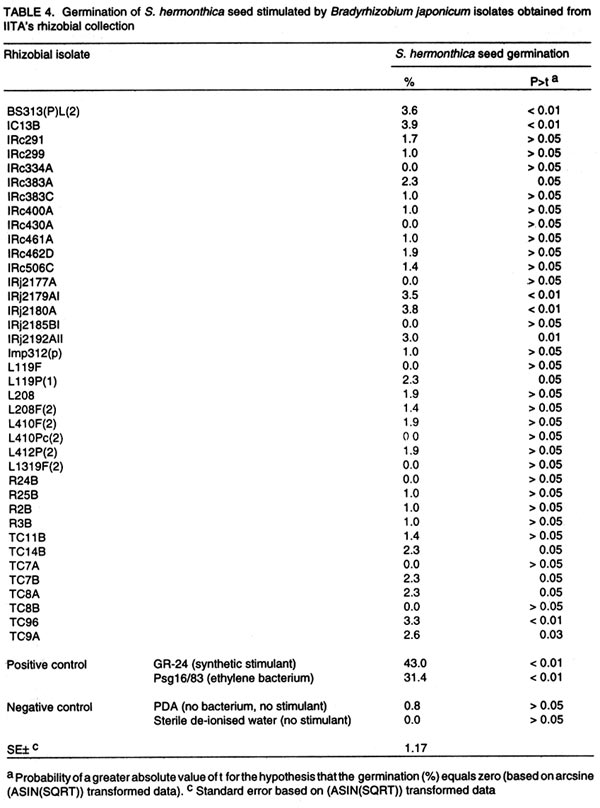
Oyo Road, Ibadan, Nigeria (Received 28 August, 2001; accepted 30 January, 2002) Code Number: cs02014 ABSTRACT Three Pseudomonas syringae pathovar glycinea (Psg) strains known to produce large amounts of ethylene were tested for efficacy in germinating seeds of Striga hermonthica (Del.) Benth. in a laboratory, as were 312 rhizosphere isolates of non-pathogenic fluorescent Pseudomonas spp. and 38 Bradyrhizobium japonicum isolates. Strains of Psg were 67% as effective as GR-24, and 31 and 38% more effective than ethylene gas (2.5 mg per Petri dish) and root pieces of the best soybean cultivar (SAMSOY-2), respectively. None of the non-pathogenic pseudomonas and rhizobial isolates effectively germinated S. hermonthica seeds compared with Psg strains. Our results indicate that ethylene producing bacterial strains can be highly effective in germinating S. hermonthica seeds and suggest that such bacteria have potential as bio-control agents of this parasitic plant in Africa. However, ethylene production by bacteria may be linked with pathogenic ability. This suggests that gene modification and transfer technology may be necessary to exploit the potential usefulness of ethylene-producing bacteria for S. hermonthica control. Key Words: Biological control, Bradyrhizobium japonicum, Pseudomonas syringae, rhizosphere RÉSUMÉ Trois souches de Pseudomonas syringae pathovar glycinea (Psg), connues pour leur capacité de produire l'ethylene en grande quantité, ont été etudiées en laboratiore afin d'evaluer leur efficacite de germer les grains de Striga hermonthica (Del.). Trois cent douze isolats rhizosphaeriques non-pathogene de Pseudomonas spp fluorescent et 38 isolats de Bradyrhizobium japonicum etaient egalement evalués. Les isolats de Psg etaient 67% aussi effective que GR-24 , et 31 et 38% plus effective que la gase ethylene (2.5 mg / Petri plat) et les pieces racinaire de la meilleure variete de soja (SAMSOY-2), respectivement. Aucun des isolats de pseudomonas non-pathogene et des isolats rhizobien n'a été effectif à induire la germinaison des grains de Striga hermonthica en comparaison avec les isolats de Psg. Ces resultats indiquent que les isolats bacterien produisant l'ethylene peuvent être hautement effective pour la germinaison des semences de S. hermonthica et suggèrent que de tel bacteries pourraient potentielement jouer un role important comme agents de lutte biologique contre cette plante parasitaire d'Afrique. Cependant, la production d'ethylene semble être liées génétiquement au pouvoir pathogène des isolats. Ceci suggère que la modification du gene et la technologie de transfer du gene pourraient être necessaire dans l'expoitation de telles bacteries produisant l'ethylene comme agent potentiel pour la lutte biologique contre S. hermonthica. Mots Clés: Lutte Biologique, Bradyrhizobium japonicum, Pseudomonas syringae, rhizosphaerique INTRODUCTION Striga hermonthica (Del.) Benth. is the most important species of the obligate root-parasitic flowering plant genus, Striga, and the major biotic constraint to cereal production in sub-Saharan Africa (Musselman, 1987). Effective control of Striga spp. is difficult mainly because of their inherently enormous reproductive ability, and high seed longevity in soil. In a single growing season, each Striga plant is capable of producing 10,000 - 500,000 seeds (Saunders, 1933), which may remain viable for up to 14 years in the soil (Saunders, 1933; Bebawi et al., 1984). If adequately conditioned, the seed would germinate only when an exogenous stimulant initiates ethylene production within the Striga seed (Vallance, 1950; Logan and Stewart, 1991; Babiker et al., 1993), or directly provides ethylene (Eplee, 1975). Since S. hermonthica is an obligate root parasite of cereal crops, stimulating germination of the parasite seeds in the absence of roots of a host cereal crop leads to death of the parasite seedlings within 3-5 days (Worsham, 1987). Such "suicidal germination" depletes the parasite's seed population in the soil and, consequently, reduces parasitism on subsequent cereal crops. Several methods for depleting Striga spp. seed bank have been used or proposed. One proven method is soil injection with ethylene gas, which has been extensively used in an eradication programme for S. asiatica in two southern states of U. S. A. (Eplee, 1975; Sand and Manley, 1990). However, ethylene gas is hazardous because it is pressurised and flammable, hence, it requires specialised storage and application equipment and technical skills for soil injection. It is costly, and therefore unsuitable in African agricultural systems. Synthetic analogues of Striga germination stimulants (Cook et al., 1972) were suggested over two decades ago for use in Striga-infested soil to stimulate suicidal germination of the parasite's seeds (Johnson et al., 1976). However, the cost, and more importantly, the instability of these analogues in soil precludes their usefulness (Babiker et al., 1987). A more practicable control option presently available to resource-poor African farmers is cereal rotation with non-host legume cultivars selected for ability to induce germination of the parasite seeds (Doggett, 1984; Berner et al., 1996a). Rotating N2-fixing grain legumes with cereal hosts can be an effective means of reducing S. hermonthica seed bank in the soil, while producing food and improving soil fertility (Berner et al., 1996b). This method is, however, very slow compared with ethylene gas, and may require several years of cultivation of selected non-host cultivars to effectively reduce parasite's seed reserves in heavily infested field soils (Eplee, 1975; Doggett, 1984; Carsky et al., 2000). Ethylene is a hormone that is produced by plants (Adams and Yang, 1979) and some microorganisms (Freebairn and Buddenhagen, 1964; Bonn et al., 1975; Goto et al., 1985; Fukuda et al., 1986; Sato et al., 1987; Nagahama et al., 1994; Weingart and Völksch, 1997). Ethylene production in natural soil has also been reported and attributed to microbial activities in soil (Kuiper, 1997; Belder, 1999). The phaseolicola and glycinea pathovars of Pseudomonas syringae produce large quantities of ethylene (Sato et al., 1987; Nagahama et al., 1994; Weingart and Völksch, 1997) which may be sufficient to induce suicidal germination of S. hermonthica. Non-pathogenic rhizobacterial strains with ability to produce effective levels of ethylene could complement cereal crop rotation with N2-fixing legume cultivars efficacious in inducing suicidal germination of S. hermonthica seeds in soil. The objectives of this study were to test the ability of known ethylene producing strains of P. syringae pv. glycinea to lead to germination of S. hermonthica seeds, and identify rhizobacterial strains effective in inducing suicidal germination of the parasite's seeds from amongst non-pathogenic groups. MATERIALS AND METHODS Germination stimulants. The P. syringae pv. glycinea (Psg) strains- 19/84, 16/83, and 8/83 (Weingart and Völksch, 1997), isolated from soybean were obtained from the Friedrich-Schiller-Universität, Jena in Germany. These were kept in 15 glycerol, and sterile water under deep freeze at -85°C, and refrigeration at the International Institute of Tropical Agriculture (IITA) Ibadan, Nigeria. They were routinely cultured on KB medium (King et al., 1954) to maintain viability. Synthetic germination stimulant, strigol, 'GR-24' (bis-lactone (XXIII)) (Johnson et al., 1976) was obtained from the NSR-Center, University of Nijmegen, Toernooeveld, The Netherlands. By dissolving 100 mg of GR-24 in 10 ml of acetone and then diluting with sterile distilled water, a 1-litre stock solution (100 mg L-1) was made and kept refrigerated. From the 100mg L-1 stock solution, a 10-mg L-1 solution was made each day of S. hermonthica seed germination testing. Ethylene gas was obtained in 13.6-kg amounts, in pressurised tanks from Machine and Welding Supply Co., Wilmington, NC. Cowpea cultivars (IT93K-637-1 and IT1137-1D) and soybean cultivars (TGX1649-11F, TGX1485-1D, and SAMSOY-2) were collected from the Grain Legume Programme of IITA. Seedlings of cowpea and soybean cultivars were raised in steam-sterilised sand in 250-ml plastic cups (three seedlings per cup) in a screenhouse at IITA. Striga hermonthica seeds and conditioning. Striga seeds used in this study were collected from S. hermonthica plants attacking sorghum (Sorghum bicolor L.), in farmers' field in Bida (latitude 9 °05 ' N, longitude 6 o 01 ' E), Nigeria in November 1997. Seeds were stored at ambient temperature in polyethylene containers, in a laboratory at IITA. To obtain conditioned S. hermonthica seeds for each day of germination testing, the S. hermonthica seeds were surface sterilised for 10 min. in an aqueous 1% NaOCl solution. Floating seeds were discarded and the remaining seeds were air-dried aseptically (under laminar flow). Moistened 3.5-mm-diameter disks of glass micro-fibre filters (GF/C, Whatman® Whatman Int. Ltd, Maidstone, England), were placed on two moistened Whatman no.2 filter papers (Whatman® Whatman Int. Ltd, Maidstone, England) in a Petri-dish, dried and then autoclaved for 15 min. at 121° C. Sterile de-ionised water (3 ml per Petri dish) was introduced to re-moisten the glass micro-fibre disks. The surface sterilised Striga seeds were aseptically placed on the sterile glass micro-fibre disks in Petri-dish as at approximately 30 seeds per disk. The dishes were placed in an incubator at 28°C, in darkness, for 14 days. To ensure that the disks remained moist, the Petri-dishes were sealed with parafilm M (Parafilm® American National Can Co. Neenah, WI 54956). Non-pathogenic rhizobacterial isolates. Three hundred and twelve isolates of saprophytic fluorescent Pseudomonas sp., were isolated from grass weed rhizosphere soils collected from four locations (Ibadan, Mokwa, Bida and Abuja) in Nigeria. In addition, 38 Bradyrhizobium japonicum isolates were obtained from the IITA's rhizobial collection (Sanginga et al., 2000). Both Pseudomonas and Bradyrhizobium isolates were kept in sterile water and PDA slants under refrigeration at IITA, and were routinely cultured on KB medium and yeast maltose agar, respectively. Testing ethylene producing P. syringae pv. glycinea strains for capacity to stimulate germination of S. hermonthica seeds. Ethylene producing P. syringae pv. glycinea (Psg) strains were tested for ability to stimulate germination of S. hermonthica seeds in Petri-dishes compared with GR-24, ethylene gas, and root pieces of two cowpea and three soybean cultivars. The experiment was conducted in a laboratory at IITA, adapting the cut-root assay (Berner et al., 1996a) to quantify stimulation of S. hermonthica seed germination by bacterial isolates, cowpea and soybean roots, ethylene gas, and GR-24. On each testing day, Petri-dishes (9-cm diameter) were lined with fresh filter papers (Whatman No. 2). A 2-cm diameter aluminium foil ring (for cut roots of soybean or cowpea), or the bottom half of an empty polystyrene 35-mm photographic film container (for bacteria and ethylene gas) was placed at the centre of each plate. The plates were autoclaved for 15 min. at 121°C, along with de-ionised water. Under a laminar flow hood (ENVIRCO, Environmental Air Control, Inc., USA), the filter papers were moistened with sterile de-ionised water (2.5 ml per Petri-dish), and the previously conditioned S. hermonthica seeds on glass micro-fibre disks, were placed around the aluminium foil ring or film container. The disks were arranged in four lines (radii), each line having four disks, with the first disk on each line, one disk-space distance away from the ring or container, and each disk touching the subsequent one. This arrangement was intended to circumvent the possible inhibitory effect on the germination of Striga seeds on the adjacent disk to the aluminium ring in the cut-root assay (Dr. Umba di Umba, IITA, Pers. comm.). Other film containers were previously autoclaved and filled with 10 ml of autoclaved potato dextrose agar (PDA). This was then allowed to solidify under aseptic conditions. A single bacterial strain was streaked onto the PDA in each of the film containers from 48-hr old bacterial culture on KB medium. The inoculated containers were then substituted aseptically for the empty containers in the centre of the Petri-dishes. For ethylene gas, balloons were used to take approximately 2.5-mg ethylene gas from pressurised tank (gas cylinder) described previously; tied and then placed in empty film containers in the centre of the Petri-dishes. Small tacks (5/8" nail) had been previously glued to the Petri-dish covers so that the tacks would be positioned over the centre of the film containers when the covers were in place. The Petri-dish covers were then pressed into position, forcing the balloons to be punctured and the ethylene gas released. The plates were immediately sealed with parafilm. To evaluate germination induction of S. hermonthica seeds by stimulants from cut-roots, root pieces of the respective crop (cowpea and soybean) cultivars were obtained from two-week old seedlings. The plants were gently removed from the cup, and washed with sterile de-ionised water to free roots of sand. The roots were then cut into 1-cm long pieces, and 1 g weighed into the aluminium foil ring in other Petri-dishes. To diffuse the root exudate across the filter paper, 300 µl of sterile de-ionised water was added to the root pieces in the aluminium foil ring. In other Petri-dishes, 300 µl of either GR-24 solution (10-mg l-1) or sterile de-ionised water was added into the central aluminium foil ring. All the Petri-dishes were sealed with parafilm immediately after introducing the stimulant. All the Petri-dishes were incubated at 33°C for 72hr after which proportions of germinated seeds were evaluated for each disk under a low power microscope. There were three experimental runs (tests), from November 1998 - January 1999, January - March 1999, and March - May 1999, each with three replications (taking a Petri-dish as replicate) per treatment. Screening of non-pathogenic rhizobacterial strains for capacity to stimulate germination of S. hermonthica seeds. Using the above procedure, 350 non-pathogenic rhizobacterial isolates made up of 312 saprophytic fluorescent Pseudomonas sp. isolates and 38 B. japonicum isolates, were screened in the laboratory. For a fast screen, there was only one day of testing for each isolate, and 12 isolates were screened per day (in one experimental run) each replicated three times. Each experimental run included 300 µl of 10-mg l-1 GR-24, and Psg strain 16/83 as positive checks, and uninoculated PDA and sterile de-ionised water as negative checks for stimulation of S. hermonthica seed germination. All the Petri-dishes were sealed with parafilm immediately after introducing the stimulant, and incubated at 33°C for 72 hr after which proportions of germinated seeds were determined under a low power microscope. Statistical analyses. All data collected were analysed using the Mixed Model procedure in SAS (Littell et al., 1996). The total number of germinated seeds and the consequent germination percentage were determined for each plate (replication). For statistical test of differences among means, germination percentage from replications were first subjected to arcsine transformation (ASIN(SQRT)), taking zero values as 10-6. For data obtained from comparing Psg strains with other germination stimulants, each experimental run was analysed by taking replication and the various germination stimulants as random and fixed effects, respectively, while in the combined analysis, experimental run and replication were both taken as random. Data from the screening of non-pathogenic rhizobacteria in laboratory were analysed by merging the separate data from each experimental run, and then taking replication and isolate as random and fixed effects, respectively. RESULTS Testing ethylene producing P. syringae pv. glycinea strains for capacity to stimulate germination of S. hermonthica seeds. Germination percentage of S. hermonthica seeds varied among germination stimulants and experimental runs (Table 1). Germination percentage of S. hermonthica seeds stimulated by each of the three Psg strains (19/84, 16/83, 8/83) were not significantly different (Table 1). Stimulation of germination was highest in the January-March 1999 test and lowest in the March-May 1999 test (Table 1). In two out of three runs, germination induced by the synthetic stimulant GR-24 was significantly higher (P<0.001) than those stimulated by Psg strains (Table 2). The Psg strains consistently led to higher germination than ethylene gas (2.5 mg ethylene per Petri-dish) or cut-root exudates of the soybean and cowpea cultivars, except for cowpea cv. IT93K-637-1 which was not significantly different from Psg strains (Table 2). Overall, mean germination stimulated by GR-24 (48.6) was highest, followed by that due to Psg (31.7), which was higher than that of ethylene gas and root pieces of cowpea and soybean cultivars, except cowpea cv. IT93K-637-1 (Fig. 1). Though some germinated S. hermonthica seeds were sometimes recorded in the no stimulant checks (PDA and sterile de-ionised water), mean germination percentages were not significantly (P<0.05) greater than zero. Screening of non-pathogenic rhizobacterial strains for capacity to stimulate germination of S. hermonthica seeds. Out of the 312 saprophytic fluorescent Pseudomonas sp. isolates screened, only 11 were found to significantly (P< 0.05) induce germination to levels greater than zero (Table 3). However, S. hermonthica seed germination by any of these isolates was not significantly greater than that on PDA without bacterial inoculation (control) (Table 3). The ethylene producing Psg strain 16/83 consistently stimulated higher (P<0.01) seed germination throughout the screening period than that on PDA with no bacterial inoculation. Results of B. japonicum isolates screening for germination of S. hermonthica seeds (Table 4), were similar to those obtained for saprophytic rhizosphere fluorescent pseudomonad isolates. Eleven of the 38 rhizobial isolates stimulated seed germination (P<0.05) to greater than zero percent (Table 4). The differences between seed germination from these isolates and those on the control, PDA with no bacterium inoculated were, however, not significant. DISCUSSION Pseudomonas syringae pv. glycinea, strains 19/84, 16/83, and 8/83, tested against other germination stimulants in this study, induced appreciable level of S. hermonthica seed germination relative to GR-24, ethylene gas and exudates from cut roots of cowpea and soybean (Fig. 1). The bacterial strains were better than 2.5 mg ethylene gas, or cut roots of three soybean cultivars and one cowpea cultivar. However, this was not true for the cut roots of cowpea cultivar IT93K-637-1, which stimulated seed germination to the same extent as the bacterial strains. As expected, GR-24 gave the highest germination stimulation. Berner et al. (1999) have also reported the same strains of Psg as effective stimulants of S. hermonthica seed germination. Germination percentage, compared with GR-24 reported by them, were not as high as in the present study. Furthermore, Berner et al. (1999), found Psg strains to induce germination of S. aspera and S. gesnerioides seeds at same levels as synthetic germination stimulant. Contrary to expectation, and Berner et al. (1999) report, a few germinated seeds were occasionally observed in the no-stimulant checks (PDA without bacterium and sterile de-ionised water). This phenomenon occurred throughout the period of the study but was negligible. It is possible that some microorganisms borne on S. hermonthica seeds produced some germination stimulants (probably ethylene) that induced S. hermonthica seed germination in spite of surface sterilisation. Another possibility is that stimulation of ethylene production within the S. hermonthica seeds through oxidation of ethylene precursor (1-aminocyclopropane-1-carboxic acid) by CO2 accumulation in the sealed plates, resulted in the germination of a few seeds without the intervention of external stimulant (Babiker et al., 2000). Ethylene gas (2.5 mg per Petri-dish) would have been expected to give a higher germination percentage than was obtained in this study. Based on surface area of the Petri-dishes, the 2.5 mg of ethylene applied per dish is equivalent to about 4 kg of ethylene ha-1. This rate is about two and half times greater than the recommended rate (1.68 kg ha-1) for S. asiatica control in United States farm fields (Eplee, 1975). Despite this large amount, the Psg strains stimulated more seed germination than did direct application of ethylene gas. This may have resulted from loss of directly supplied ethylene from the Petri-dish (and a lower concentration than intended) in the interval between puncturing the gas-filled balloon and sealing the Petri dish with parafilm. On the other hand, the growing bacteria released ethylene on a continuous basis within the sealed Petri-dishes, hence, was more effective in sustaining germination stimulation over time. Another possibility could be that extremely high concentrations of exogenous ethylene might partially retard germination, or that seeds of S. hermonthica respond differently to different ethylene concentrations compared to those of S. asiatica. In this regard, Babiker et al. (1993) found that seeds of S. asiatica germinated, in response to 0.8 - 1.0 µl l-1 of ethylene while Jackson and Parker (1991) found that S. forbesii Benth. seeds were non-responsive to 1.0 µl l-1 exogenous ethylene. The Psg strains; 16/83 and 19/84 produced ethylene at about 95 X 10-8 nl h-1 cell-1, while strain 8/83 produced ethylene at about 25 X 10-8 nl h-1 cell-1 (Weingart and Völksch, 1997). Despite the lower ethylene production, strain 8/83 induced similar levels of germination as the other two strains (Table 1), indicating that the bacteria strains produce more ethylene than the threshold required for germination by S. hermonthica seeds. Cowpea and soybean cultivars (non-hosts of S. hermonthica) selected for their efficacy in stimulating germination of S. hermonthica have been used in field studies to reduce parasitism on sorghum and maize (Doggett, 1984; Ariga, 1996; Berner et al., 1996a, b; Carsky et al., 2000). The roots of such non-hosts putatively produce germination stimulants that induce endogenous ethylene production within Striga spp. seeds (Logan and Stewart, 1991; Babiker et al., 1993); the seeds then germinate in absence of the host and, therefore, die within some days. Rotation of host cereal crops with these non-host germination-stimulant producers reduces the amount of parasite seeds in soil and, therefore, the level of parasitism on subsequent host cereal. To be acceptable for use in agricultural soils, the ethylene-producing bacteria must be proved non-pathogenic to crop plants. Pseudomonas syringae pathovars are well known pathogens of important crop plants and are, therefore, unacceptable for use as biological control agents in African agricultural soils. In this study, 312 non-pathogenic species of fluorescent Pseudo-monas sp. were screened in search of non-pathogenic ethylene producing rhizobacterial strains capable of stimulating germination of S. hermonthica seeds at levels comparable to those of Psg strains. In addition, 38 B. japonicum isolates from soybean and cowpea root nodules were screened. Eleven isolates each of Pseudomonas sp. and B. japonicum induced germination of S. hermonthica seeds to levels significantly (P<0.05) greater than zero. However, none of the germination levels obtained from these bacterial isolates was significantly better than the rare germination in the no-stimulant checks. It is possible that none of these bacterial isolates produce ethylene or that they produce ethylene at levels too low to give high levels of stimulation of S. hermonthica seeds germination. Plant pathogens are reported to be efficient ethylene producing micro-organisms (Freebairn and Buddenhagen, 1964; Bonn et al., 1975; Pegg and Cronshaw, 1976; Goto et al., 1985; Fukuda et al., 1986; Sato et al., 1987; Nagahama et al., 1994; Weingart and Völksch, 1997). As suggested by some workers (Pegg and Cronshow, 1976; Nagahama et al., 1994), a high level of ethylene production in micro-organisms may be a trait linked to plant pathogenesis. If this is true, then the only option left to exploit this native microbial property of ethylene production for S. hermonthica biological control would be the use of gene technology. Fortunately, the genes for ethylene production in the glycinea and phaseolicola pathovars of P. syringae are carried on plasmids of the bacterium (Nagahama et al., 1994). Therefore, the plasmids could be genetically modified and transferred into non-pathogenic soil-borne rhizosphere fluorescent Pseudomonas species that could then be used as biocontrol agent of S. hermonthica in agricultural soils, through suicidal germination of the parasites' seeds. Non-pathogenic ethylene producing or ethylene-forming-enzyme gene-bearing rhizobacterial strains can be inoculated to seeds of cowpea and soybean cultivars (non-hosts of S. hermonthica), in rotation with cereals. By combining such bacteria with non-host crops, effective in inducing germination of S. hermonthica seeds such as the cowpea cv. IT93K-637-1 and soybean cv. SAMSOY-2 tested in this study, it may be possible to increase suicidal germination of the parasites' seeds compared with non-host crop rotation alone. Plant growth promoting rhizosphere Pseudomonas spp. strains (PGPRP) with efficient ethylene producing capacity would, particularly be useful in this respect because of their nutritional versatility and fast growth rate and, therefore, establishment in rhizosphere of many crops (Kleopper et al., 1980; Weller, 1983). Besides, B. japonicum strains effective in cowpea and soybean root nodulation (N2-fixation) are available (Sanginga et al., 1996; 2000) that could improve soil N2 fertility capable of reducing S. hermonthica parasitism in subsequent cereal crop. Research aimed at co-inoculating seeds of cowpea and soybean cultivars selected for suicidal germination of S. hermonthica with efficient ethylene producing PGPRP and B. japonicum is, therefore, desirable. ACKNOWLEDGEMENTS The authors are grateful to the anonymous reviewers selected by IITA Publications Review Panel for their useful comments. This paper is approved for publication as International Institute of Tropical Agriculture manuscript number: IITA/01/JA/24. REFERENCES
©2002, African Crop Science Society The following images related to this document are available:Photo images[cs02014t1.jpg] [cs02014t4.jpg] [cs02014t2.jpg] [cs02014f1.jpg] [cs02014t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}