 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
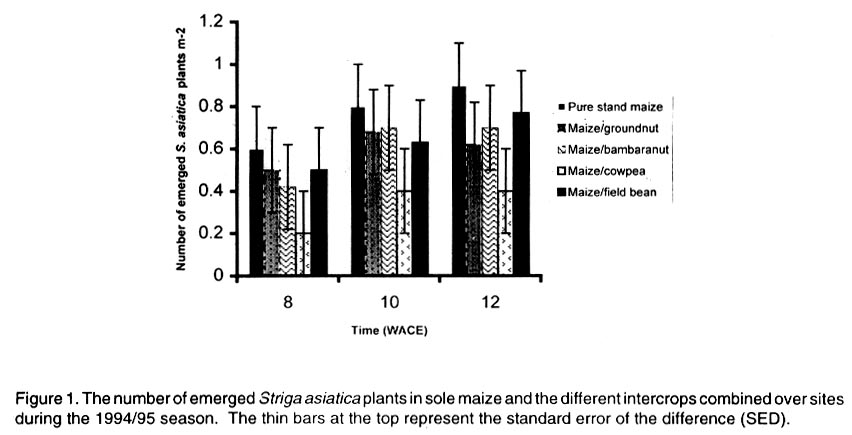
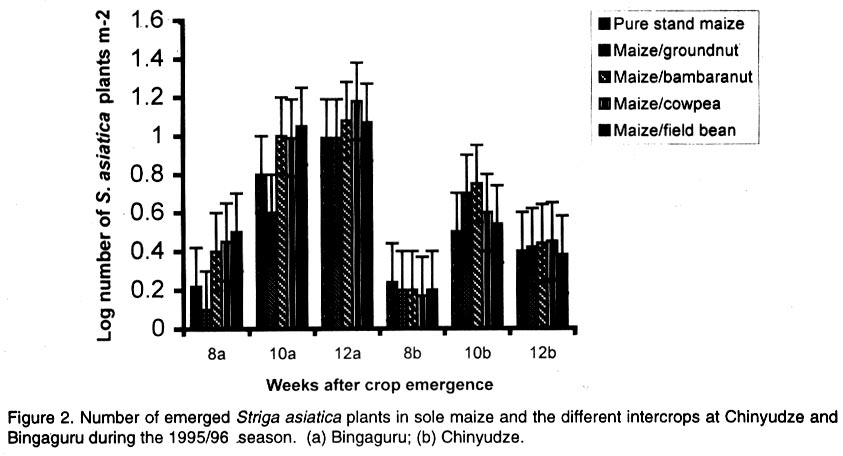
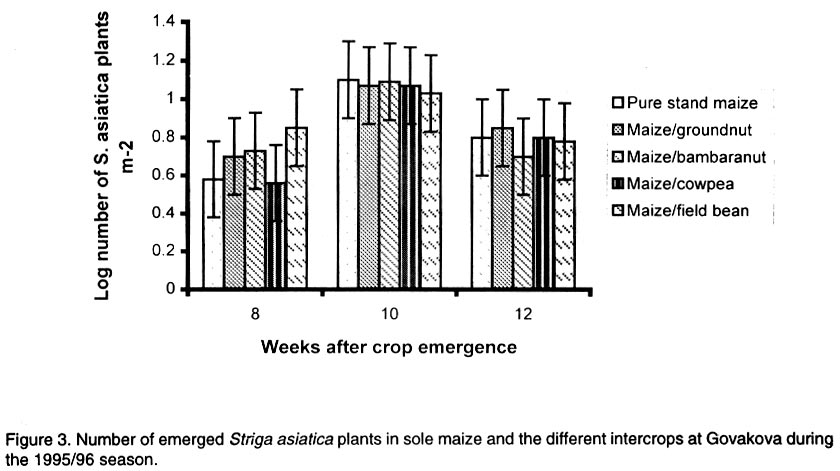
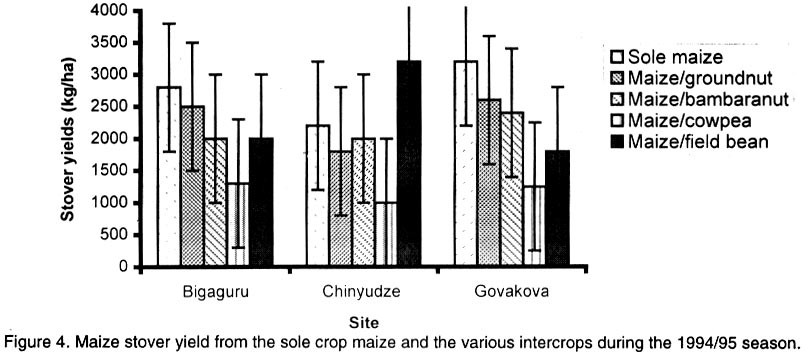
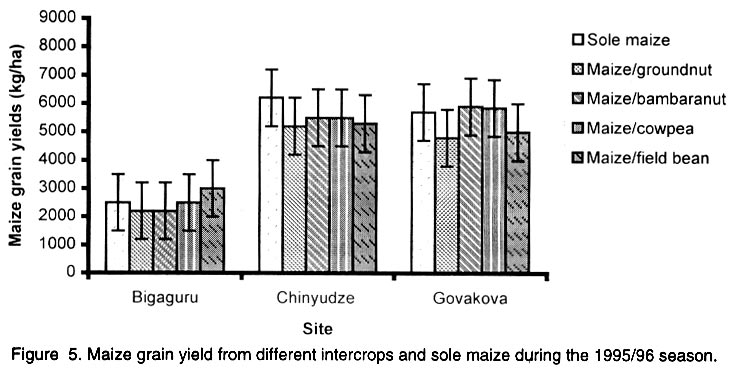
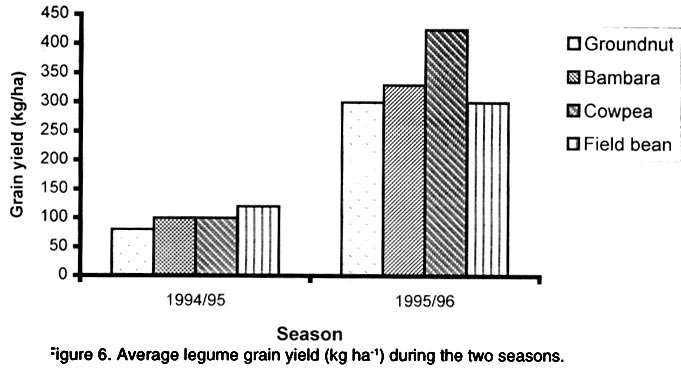
African Crop Science Journal, Vol. 10. No. 2, 2002, pp. 163-171 Intercropping maize with grain legumes for Striga control in Zimbabwe D. Musambasi, O.A. Chivinge and I.K. Mariga University of Zimbabwe, Crop Science Department, P.O. Box MP167, Mount Pleasant, Harare, Zimbabwe (Received 9 March, 2001; accepted 10 December, 2001) Code Number: cs02016 ABSTRACT On-farm experiments were conducted in Chinyika Resettlement Area during the 1994/95 and 1995/96 rainy seasons at four sites to test the effectiveness of intercropping maize with cowpea, groundnut, field bean and bambara nut in suppressing Striga asiatica (L.) Kuntze. During the 1994/95 rainy season maize/cowpea intercrop supported less S. asiatica plant m-2 than sole maize, while other intercrops supported similar numbers of emerged S. asiatica plants m-2 as sole maize. Combined analysis over sites showed that the numbers of emerged S. asiatica plants in the intercrops were similar to those in sole maize during the 1995/96 season. However, during the same season the number of emerged S. asiatica plants was significantly (P<0.05) lower in the maize/groundnut intercrop at 8 weeks after crop emergence (WACE) at one site but similar at the remaining sites. Due to moisture stress and severe competition from the parasitic weed, no maize grain was harvested during the 1994/95 season. The maize/cowpea intercrop had the lowest maize stover yield, while the maize/field bean intercrop had the highest stover yield. Maize grain yields were not significantly different for all treatments during the 1995/96 season but the partial land equivalent ratios (PLERs) ranged from 0.8 to 1.70 indicating that intercropping maize with legumes had an advantage over planting sole maize in S. asiatica infested fields. Key Words: Competition, moisture stress, partial land equivalent ratio, Striga asiatica, yields Résumé Des expériences sur fermes ont été conduites, en quatre endroits du camp de réinstallation de Chinyika, pendant les saisons 1994/1995 et 1995/1996 pour tester l'efficacité de la culture du maïs et du niebé intercalé, l'arachide, l'haricot et l'arachide bambara a supprimé le Striga asiatica (L.) Kuntze. Pendant la saison 1994/1995 le maïs combiné au niebé a supporté moins de Striga asiatica que le maïs en monoculture, tandis que les autres cultures associées ont supporté une émergence identique du striga asiatica comme le maïs en monoculture. L'analyse combinée des sites a montré que l'émergence du striga asiatica dans les cultures combinées était similaire à celle du maïs durant la saison 1995/1996. Durant la même saison cette émergence était significativement faible dans le maïs combiné à l'arachide 8 semaines après l'émergence de la plante dans un site seulement. A cause du déficit en eau et une sévère compétition de la part des parasites aucune graine de maïs n'a pu être récoltée durant la saison 1994/1995. La culture du maïs combine au niebé avait le plus faible rendement en biomasse de maïs; alors que le maïs combiné à l'haricot a produit le plus grand rendement. Les différences de rendement n'ont pas été significatives pour tous les traitements durant la période 1995/1996. Le taux partiel de terres équivalentes a range entre 0.8 et 1.70 indiquant que la culture du maïs combiné aux légumes a eu un avantage sur la culture du maïs seul concernant l'incidence du S. asiatica. Mots Clés: Compétition, stress lié à l'humidité, taux partiel de terres équivalentes, Striga asiatica, rendements INTRODUCTION Striga asiatica (L.) Kuntze (witchweed) is a weed which causes 50-100% decrease in grain yields of cereal crops (Berner et al., 1995; Musambasi, 1997). The weed is prolific, and produces between 50 000 and 500 000 seeds plant-1 which can remain viable up to 20 years (Berner et al., 1995). An effective Striga management programme should include a reduction in the number of Striga seed from the heavily infested soils and prevention of further seed multiplication (Ramaiah, 1983). Rotating or intercropping maize with trap crops such as field beans (Phaseolus vulgaris L.), groundnut (Arachis hypogea L.), bambara nut [Vigna subterranea (L.) Walp], sunflower (Helianthus annus L.) and cowpea [Vigna unguiculata (L.) Walp] may help to reduce the number of S. asiatica seed in the soil. Farmers in Chinyika Resettlement Area already intercrop maize with some of these legumes, which have added advantages of being bonus crops, adding nitrogen to the soil and suppressing the germination and growth of other weed species through shading. The trap crops such as cowpea induce S. asiatica seed germination but do not support its subsequent growth and development. In the absence of a suitable host the S. asiatica seedling dies within four days from germination (Butler, 1989; Ejeta and Butler, 1993). Therefore, trap crops offer excellent scope to control this weed because they not only reduce the S. asiatica seed reservoir but also enhance soil fertility through N fixation and thus lead to increased grain yields of subsequent cereal crops (Odhiambo and Ransom, 1994). Carson (1989) showed that intra-row intercropping of sorghum [Sorghum bicolor (L.) Moench] and groundnut significantly reduced Striga hermonthica (Del) Benth emergence. This was associated with a decrease in soil temperature in the intercropped plots. In Gambia, alternating sorghum or millet [Pennisetum americanum (L.) K. Schum] with groundnut resulted in a low S. hermonthica infestation (Lagoke et al., 1988). Soybeans (Glycine max L.), cotton (Gossypium hirsutum L.) and bambara nut when grown as intercrops with maize or sorghum, are known to induce abortive germination of S. hermonthica seeds, with a consequent reduction in infestation. This has advantages of depleting the seedbank and ensuring that no new seed is added to the soil (Sauerborn, 1999). The objectives of this study was, therefore, to determine the effects of interrow intercropping of groundnut, field bean, cowpea and bambara nuts with maize on S. asiatica and intercrop performance. MATERIALS AND METHODS The experiment was conducted on-farm in Chinyika Resettlement Area (32° 05" and 32° 44" E and 18° 00" and 18° 05" S) in the eastern part of Zimbabwe, during the 1994/95 and 1995/96 rainy seasons. Four sites (two in Chinyudze and one each in Govakova and Bingaguru) were used in both seasons. Chinyudze and Bingaguru are in Natural Farming Region (NFR) IIb (750-1000 mm annum-1 with mid-season dry spells), while Govakova is in NFR III (600-800 mm annum-1). Farmers ploughed the fields using ox-drawn ploughs in early October of each year, while pegging of field plots and opening furrows were done just before the onset of the rains in mid to late October. Immediately after the first effective rains, the four legumes (local landraces of groundnut, bambara nut, cowpea and field beans) were simultaneously intercropped with maize hybrid SC501, while a sole maize stand was used as a control treatment. Compound D (8% N: 14% P2O5: 7% K2O) was applied as basal fertiliser at planting at a rate of 300 and 200 kg ha-1 for maize and legumes, respectively. Maize was topdressed 5 weeks after crop emergence (WACE) with 250 kg ha-1 ammonium nitrate (34.5% N), while 200 kg ha-1 gypsum [calcium sulphate (17.5 % S)] was applied to groundnut plants at flowering. The experiment was arranged in a randomised complete block design (RCBD) with four replicates. All sites had six rows of maize per plot, 6 m by 5.4 m wide, while the legumes had 12 rows of the same length as the maize rows. The spacings were 90 cm x 30 cm for maize and 45 cm x 10 cm for the legumes giving a plant population of 37,037 and 222,222 ha-1 for maize and legumes, respectively. The gross plot area was 32.4 m2 while the net plot area was 14.4 m2 for maize and 7.2 m2 for legumes. The number of emerged S. asiatica plants was counted from the whole plot at 8, 10 and 12 WACE. These were later expressed as emerged S. asiatica plants m-2 before being subjected to logarithm transformation using the formula log (X+1), where X is the number of emerged S. asiatica plants m-2. Maize and legume grain yields were obtained from the respective net plots. Combined analysis over sites, in each season was done after performing the Bartlett Test of homogeneity of variance. RESULTS Density of Striga asiatica plants. In general the intercrops supported less S. asiatica plants that emerged than the sole maize. The highest number of S. asiatica plants emerged in sole maize, while the maize/cowpea intercrop supported the least number at all sites during the 1994/95 season (Fig. 1). The maize/cowpea intercrop supported 78.8, 54.5 and 59.1% less S. asiatica plants than the sole crop maize at 8, 10 and 12 WACE, respectively. In addition, it had 50% less S. asiatica plants than the other intercrops. Furthermore, some S. asiatica plants in the maize/cowpea intercrop appeared whitish due to what was suspected to be a fungal infection. However, during the 1995/96 season the number of S. asiatica plants that emerged were not significantly different (P<0.05) in all treatments at Chinyudze, while at Bingaguru the maize/groundnut intercrop supported the least number of S. asiatica plants that emerged at 8 WACE (Fig. 2). The maize/field bean, maize/bambara nut and the maize/cowpea supported similar numbers of the parasite at 8, 10 and 12 WACE at Bingaguru (Fig. 2). The peak period of S. asiatica plants emergence was at 10 WACE because there were more S. asiatica plants than at 8 and 12 WACE at Chinyudze and Govakova (Figs. 2 and 3). Combined analysis over sites during the 1995/96 season showed that the number of S. asiatica plants that emerged were not siginificantly different (P<0.05) in all the intercrops. Maize grain and stover yields. During the 1994/95 season, the maize/cowpea intercrop had the lowest maize stover yield, while the maize/field bean intercrop resulted in the highest yield at Chinyudze (Fig. 4). The maize/field bean intercrop outyielded the maize/cowpea intercrop by 2,042 kg ha-1 at Chinyudze during the 1994/95 season, while at Bingaguru the difference was only 700 kg ha-1 (Fig. 4). The maize/groundnut, and the maize/bambara nut intercrops stover yields were not significantly different (P<0.05) at all the sites in the same season. At Govakova and Bingaguru, the maize/cowpea and the maize/field bean intercrops had stover yields, which were lower than that from sole maize. Maize grain yields during the 1995/96 season were not significantly different (P<0.05) in all treatments at all sites, averaging 2,400 kg ha-1 at Bingaguru and 5,500 kg ha-1 at the other three sites (Fig. 5). When the data were combined over sites in the two seasons, there were no significant (P<0.05) treatment effects on maize stover yields in the 1994/95 season and maize grain yields in the 1995/96 season. Legume yields. The legumes had lower unshelled grain yields during the 1994/95 season but the yields improved considerably during the 1995/96 season, which received 884 mm year-1 , the normal amount of rainfall expected in the area. Generally the legumes had unshelled grain yields below 115 kg ha-1 during the 1994/95 season and above 293 kg ha-1 during the 1995/96 season (Fig. 6). Partial land equivalent ratios (PLER). Table 1 summarises the PLER of maize from the various intercrops for the 1995/96 season. Intercropping maize and bambara nut at Chinyudze, Govakova and Pfumoiguru, maize and groundnut at Pfumoiguru, maize and cowpea at Govakova and Pfumoiguru, and maize and field bean at Chinyudze and Pfumoiguru resulted in greater than unit (above 1) PLER's for maize (Table 1). The least PLER obtained was 0.82 at Govakova for the maize/groundnut intercrop (Table 1). DISCUSSION Density of S. asiatica plants. The lower number of S. asiatica plants that emerged in the intercrops during the 1994/95 season indicated a reduced potential for overall flower and capsule production and, consequently, a reduced capacity of increasing the S. asiatica seed bank in the soil. It is, however, important to note that the number of S. asiatica plants that emerged represents an unknown and often variable percentage of the total number of S. asiatica plants that actually parasitize the host's roots. The decreased number of S. asiatica plants in the maize/cowpea intercrop may be attributed to the suicidal germination caused by the germination stimulant produced by the cowpea roots. In addition to being a trap crop, cowpea provides shade which smothers the witchweed thereby reducing its vigour. These two factors are detrimental to the growth and development of S. asiatica plants. Striga asiatica transpires less when shaded, thereby reducing the amount of nutrients and water drawn from the maize (Stewart and Press, 1990). Fungal diseases are also prevalent when the witchweeds are shaded as observed in the maize/cowpea intercrop during the 1994/95 season. S. asiatica plants that occupy the lower vegetative stratum are adversely affected and, hence, less of the weed reaches the reproductive stage. Bebawi (1987) and Ndikawa and Kenga (1993) reported the same trend when cowpea was either intercropped with maize or sorghum. Kabambe (1995) also reported less numbers of emerged S. asiatica plants in the maize/cowpea intercrop using an intra-row intercropping pattern. Carson (1989) reported reduced numbers of emerged S. hermonthica plants when sorghum was intercropped in the same row with groundnut. The results from this study indicate that although groundnut suppresses S. asiatica plants, the suppressive effect was less in extent than for cowpea in the 1994/95 season. However, intra-row intercropping pattern may cause more S. asiatica seedlings to attach to the maize roots because of the close proximity of cowpea and maize roots. This view was also supported by Berner et al. (1995). In inter-row intercropping, the maize and cowpea roots are not close and the chances of the germinating seedlings attaching to the maize roots are less. During the 1994/95-season soil moisture was very limiting and as a result, there was intraspecific competition for moisture and attachment sites among the germinating S. asiatica seeds. Cowpea as an efficient trap crop stimulated more seeds to germinate but only a fraction survived because not all the germinated seeds could attach to the maize roots. During the 1995/96 season, which was a wetter season, most of the stimulated S. asiatica seeds germinated and a greater number could have survived and attached to the maize roots because moisture was not limiting. As a result, no significant differences in the number of S. asiatica plants that emerged were observed between the various intercrops and the sole maize crop. Germination and emergence in Striga species vary from season to season, depending on weather conditions especially soil temperature and moisture relations (Dogget, 1988). Oryokot (1993) also reported a seasonal dependence of Striga species emergence even under laboratory conditions. This could explain why S. asiatica dynamics were different in the two seasons. Many environmental factors including soil temperature and moisture status, may affect the growth and development of Striga species either acting directly on the weed or by mediation through the host (Oryokot, 1993). These factors probably affected the after-ripening period, conditioning of the seed, germination or early seedling development resulting in no differences in S. asiatica numbers obtained during the 1995/96 season. The performance of the different intercrops was not consistent across sites but, it must, however, be emphasized that there was a trend that the maize/cowpea intercrop supported less S. asiatica numbers than the other intercrops. Maize grain and stover yields. There were no maize grain yields during the 1994/95 season due to a combination of devastative effects of both S. asiatica infestation and adverse weather conditions. Striga asiatica causes more damage to the maize crop if there is limited moisture because it takes most of the water from its host. The effectiveness of S. asiatica in obtaining water from its hosts is ascribed to its leaf tissue having a greater osmotic pressure than the host leaves and to its leaves having low stomatal resistance which provides for higher transpiration rates (Stewart, 1987). This higher transpiration rate renders S. asiatica a stronger sink for the solutes and water than maize. Nutrient uptake is greater in intercrop systems than in sole crops and under low fertility systems, there is more competition from maize because it has a fibrous rooting system compared to the tap root system of legumes. However, under low rainfall conditions the legumes that have a deeper tap root system compared to maize do well. This may explain why no maize grain yields were obtained during the 1994/95 season, which was characterised by a pronounced drought period. Despite supporting the least number of S. asiatica plants, the maize/cowpea intercrop had the lowest maize stover yield mainly because interspecific competition between maize and cowpea is more pronounced under adverse moisture conditions such as those that prevailed in the 1994/95 season. This exerted further stress to the maize crop and hence failure to channel resources for reproductive growth. In addition, the duration of the competition was longer than in any other intercrop because the cowpea strain used had an indeterminate growth habit requiring a long growing season. However, there was a total crop failure at one site in the 1994/95 season as the rainfall received (485.5 mm annum-1) was well below the normal expected (650-800 mm annum-1) in average seasons. Water stress was aggravated by infestation by S. asiatica, which is a strong sink for moisture and other assimilates because of its high transpiration rates. As a result, maize was severely stressed resulting in poor pollen-silk synchronization and consequently failure to produce grain. Kabambe (1995) reported lower maize grain yields in lower rainfall areas of Malawi when maize was intercropped with cowpea for S. asiatica control. However, the maize stover yield was highest in the maize/field bean intercrop at Chinyudze during the 1994/95 season because the field bean crop took less time to mature reducing competition for water and other resources. Maize grain yields were similar in all the intercrops during the 1995/96 season and this may indicate that in a good season, the type of legume intercropped with maize does not affect maize grain yields. However, there may be differences in the beneficial effects on maize yields in subsequent seasons due to residual nitrogen, which would have been fixed by the particular legume. Under dry environments, the disadvantage of competition for moisture and other resources probably outweighs the advantage of reduced S. asiatica load and this should explain why no grain yield were realised in the 1994/95 season. CONCLUSIONS AND RECOMMENDATIONS The maize/cowpea intercrop exhibited a higher potential for suppressing S. asiatica emergence as compared to the other intercrops especially during the drier season. The maize/groundnut and maize/bambara nut intercrops suppressed S. asiatica by the same magnitude in the 1994/95 season. During the wetter season the number of emerged S. asiatica plants in the intercrops and in the sole maize crop was similar. Further investigations need to be conducted with particular emphasis on the cowpea germplasm, intercropping pattern and plant densities in the intercrop in order to come up with a sound recommendation to the farmers. In the wetter season the intercrops need to be complemented with handweeding in order to prevent the parasite from flowering and setting seeds. ACKNOWLEDGEMENTS Research was supported by Rockefeller Forum for Agricultural Husbandry in Southern and Eastern Africa and conducted in Chinyika Resettlement Area in Zimbabwe. The authors thank the technical staff at the Department of Crop Science and field assistants in Chinyika Resettlement Area for their support, and the farmers for hosting the trials. REFERENCES
©2002, African Crop Science Society The following images related to this document are available:Photo images[cs02016f5.jpg] [cs02016t1.jpg] [cs02016f3.jpg] [cs02016f2.jpg] [cs02016f4.jpg] [cs02016f1.jpg] [cs02016f6.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}