 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 10. No. 2, 2002, pp. 183-194 Regional distribution of lepidopteran stemborers and their parasitoids among wild grasses in the semi-arid eastern Kenya J. M. SONGA, W. A. OVERHOLT1, J. M. MUEKE2 and R. O. OKELLO2 National Dryland Farming Research Centre, Katumani, P. O. Box 340, Machakos,
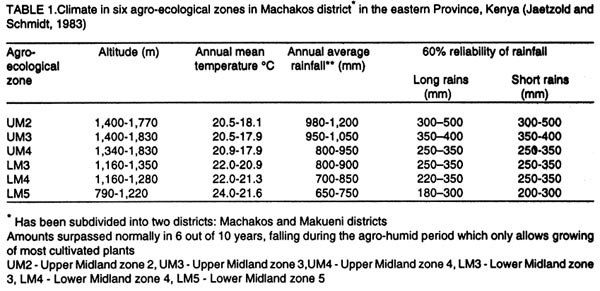
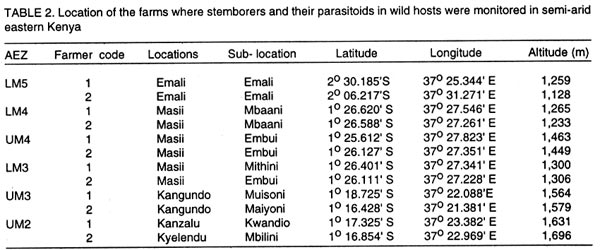
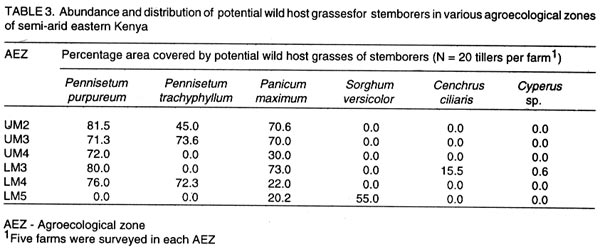
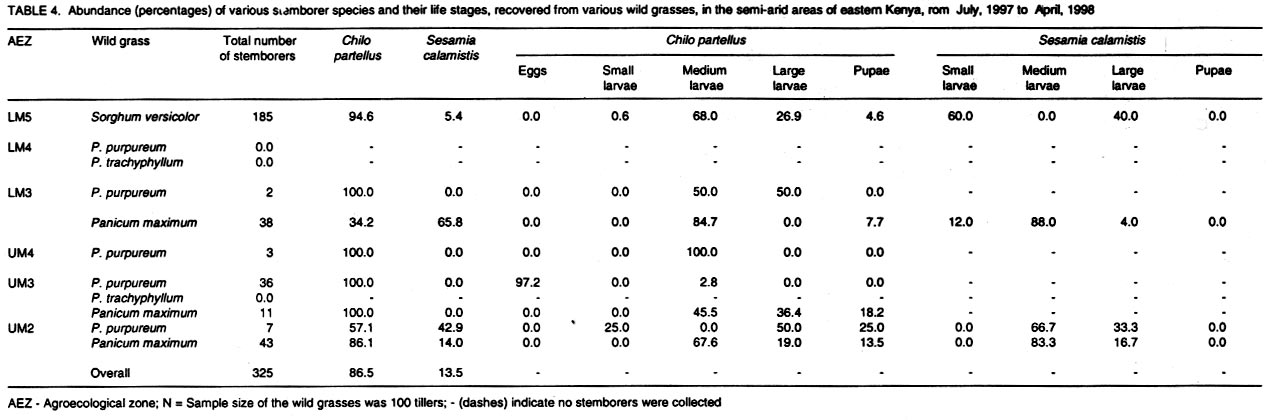
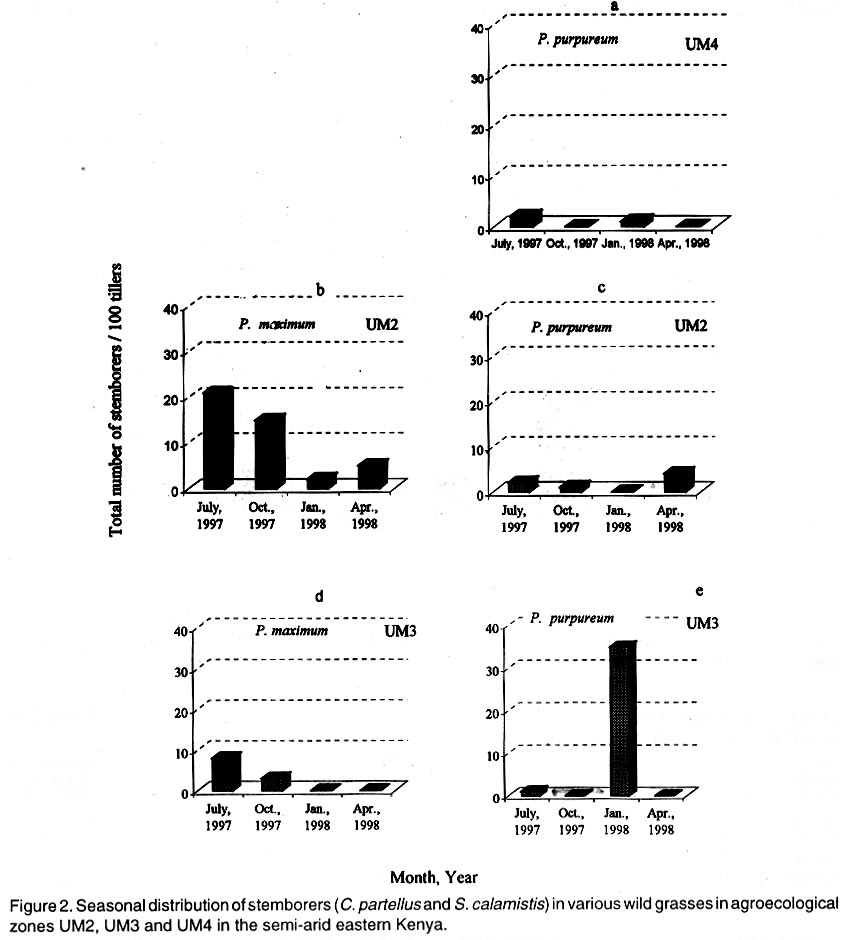
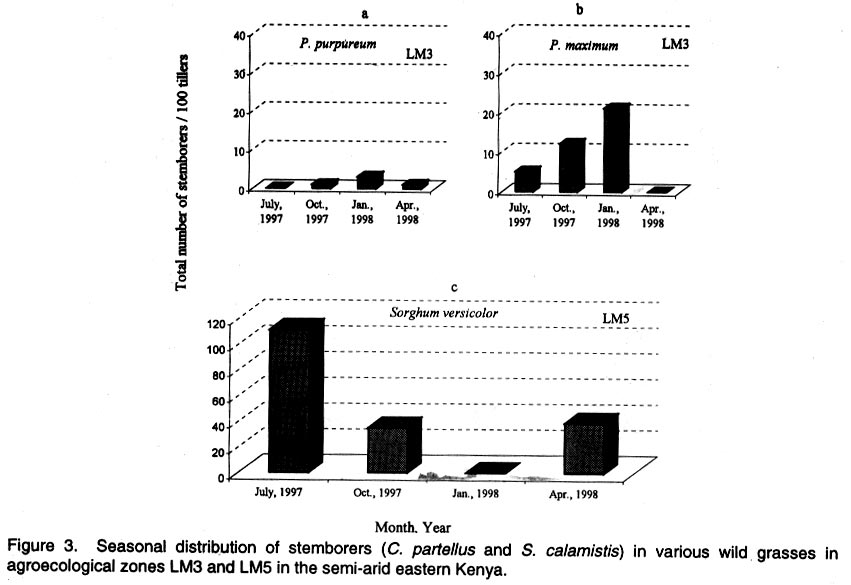
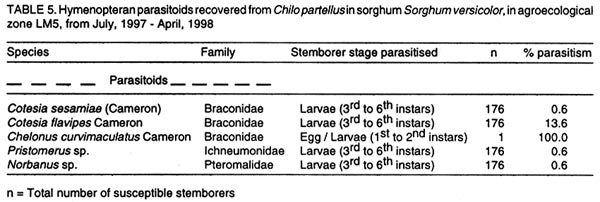
Kenya. (Received 25 April, 2001; accepted 5 March, 2002) Code Number: cs02018 ABSTRACT A survey was conducted in six agroecological zones (AEZs) in semi-arid eastern Kenya, in May, 1996, to identify the major wild host grasses (abundance > 70%) for stemborers in each zone. The grasses sampled were Pennisetum purpureum Schumach, P. trachyphyllum Pilg., Panicum maximum Jacq, Sorghum versicolor Anderss, Cenchrus ciliaris L. and Cyperus sp. Among these grasses, P. purpureumwas the most widespread and abundant. The survey was followed by an on-farm study conducted in each of the six AEZs to determine the relative abundance and distribution of stemborers, that infested the major wild grasses and their parasitoids . This was done by destructive sampling of 100 tillers, at a three-month interval, from July 1997 to April 1998. The stemborers that infested the grasses were Chilo partellus Swinhoe and Sesamia calamistis Hampson. Among these, C. partelluswas the predominant species, with the highest number being recovered on S. versicolor (94.6%). Parasitoids were only reared on S. versicolor, and these were Cotesia sesamiae (Cameron), C. flavipes Cameron, Chelonus curvimaculatus (Cameron), Pristomerus sp. and Norbanus sp., with C. flavipes causing the highest parasitism (13.6%). Results reveal that S. versicolor exhibited potential for use in the management of stemborers in maize, by acting as a "trap" and "refuge" crop for C. flavipes, between cropping seasons. Key Words: Agroecological zones, Chilo partellus, Cotesia flavipes, Cotesia sesamiae, refuge, trap crop RÉSUMÉ Une reconnaissance a été éffectuée dans six zones agro-écologiques de la région semi-aride du Kenya, en Mai 1996, en vue d'identifier les herbes sauvages dominantes (abundances > 70%) pouvant servir de pièges pour les destructeurs des tiges de maïs. Les herbes échantillonnées étaient Pennisetum purpureum Schumach, P. Tranchyphyllum Pilg., Panicum maximum Jacq, Sorghum versicolor Anderss, Cenchrus ciliaris L. and Cyperus sp. La reconnaissance était appuyée par une étude dans les fermes, dans chaque zone écologique, pour déterminer l'abondance relative et la distribution des destructeurs des tiges et leurs parasites envahissant les herbes dominantes. Pour ce faire, un échantillonnage destructif était utilisé. Cent tiges fraîches étaient coupées à trois mois d'interval, de Juillet 1997 en Avril 1998. Les destructeurs des tiges ayant attaqué les plantes étaient Chilo partellus Swinhoe et Sesamia calamistis Hampson. Partellus était l'espèce la plus dominante. Partellus était abondant sur le S. versicolor. Les parasites se cabraient uniquement sur S. versicolor, parmis eux, Cotesia sesamiae (Cameron), C. flavipes Cameron , Chalonus curvinaculatus (Cameron), Pristomerus sp. et Norbanus . Flavipes C. avait le plus taux de parasitisme (13.6%). Les résultats ont ainsi révélé que S. versicolor a un potentiel de control sur les destructeurs de mais en agissant comme piège et refuge au C. flavipes durant les saisons de culture. Mots Clés: Zones agro-écologiques, Chilo partellus, Cotesia flavipes, Cotesia sesamiae, refuge, plant piège INTRODUCTION Lepidopteran stemborers are one of the major constraints to maize production in the semi-arid areas of eastern Kenya (Darling, 1993; Songa et al., 1999). The semi-arid and arid areas cover 80% of the country and 50% of Kenya's arable land (Jaetzold and Schmidt, 1983). Most stemborers that attack maize, are polyphagous; they feed on several other cultivated Graminaceous plants and on wild host grasses. Various wild host plants of these stemborers have been documented (Ingram, 1958; Nye, 1960; Harris, 1962; Gebre-Amlak, 1988). Studies in the Kenyan Coast have shown that, during cropping seasons, stemborers occur in high numbers in maize fields. However, after harvest of the crop, some of the stemborers (adults) migrate to alternate hosts and wild host grasses close to the fields, where they survive throughout the non-cropping dry season. Some of the stemborer larvae enter into diapause in maize residues left in the field after harvest (Warui and Kuria, 1983; Unithan and Seshu Reddy, 1989). Soon after the onset of rains, diapause of the larvae is terminated, allowing development into pupae and adults that move back into the young growing crops (Scheltes, 1978) and neighboring wild grasses. Some of the stemborers from the wild grasses also reinfest the young maize plants (Mbapila, 1996). Wild host grasses have been reported to contribute to the carry-over of stemborer populations between seasons (Walker, 1983). Removal and destruction of these wild grasses has been suggested as a method for reducing stemborer populations in maize fields (Nye, 1960; Wheatley, 1961; Roome, 1976; Seshu Reddy, 1983). However, the effectiveness of this method (destruction of wild grasses) lies on application by all farmers in the target zone, which is most unlikely (Adesiyun and Ajayi, 1980; Ajayi, 1989). Alternatively, wild host grasses adjacent to maize crops, are considered to be beneficial because they provide refuge for natural enemies as well as a source of nectar, pollen, and host/alternate prey, during the non-crop periods (Altieri et al., 1977; Altieri and Whitcomb, 1979; Herzog and Funderburk, 1986; Khan et al., 1997). The presence of some wild grass species near maize crop fields may therefore contribute to reduction of stemborer infestations in the maize crop, by facilitating the carry-over of natural enemies of stemborers from one cropping season to the next. Alternate hosts and wild host grasses may also be used as trap crops during crop growth, and may reduce the pest population on the cultivated Graminaceous crops (Randriamananoro, 1996). Information on the alternate hosts of stemborers is therefore important for the development of an integrated pest management strategy for stemborers in the semi-arid eastern Province of Kenya. However, knowledge of the wild host grasses of stemborers, the stemborer complex that attacks them, and their associated parasitoids, is lacking in these areas. The objectives of this study therefore were to determine i) the relative abundance of potential wild host grasses for stemborers in semi-arid Eastern Kenya and ii) the range and distribution of stemborer species that infest these grasses and their associated parasitoid. MATERIAL AND METHODS Selection of wild grasses for the on-farm study. This was done through a preliminary survey, which was conducted in five farms in each of the six agroecological zones (AEZs): upper midland zone 2 (UM2), UM3, UM4, lower midland zone 3 (LM3), LM4 and LM5 (Table 1; Fig. 1), in the semi-arid eastern Province of Kenya, in May, 1996. The aim of the survey was to assess the relative abundance of potential wild host grasses for stemborers along field margins up to a distance of 10 m from the edge of the crop fields. Potential wild host grasses for stemborers were identified based on available information on the work carried out in Nyanza, Western (Randriamanoro, 1996) and Coast provinces of Kenya (Mbapila, 1996), and on visual symptoms of stemborer damage on the plants in the study sites. A wooden quadrant of size 1m x 1m was used to assess the percentage area occupied by each of the potential wild host grass species in each of 20 randomly selected positions per farm. The abundance and distribution of each of the wild host grasses was determined for each of the six AEZs. In each of the AEZs, all the potential wild host grasses for stemborers that had an abundance of more than 70% were selected for use in the subsequent on-farm study. Distribution of stemborers and parasitoids. This study was conducted in two farms in each of the six AEZs (UM2, UM3, UM4, LM3, LM4, LM5) in semi-arid eastern Kenya (Tables 1 and 2; Fig. 1). In each farm, two existing terrace edges (the area between the two terraces was under maize crop), along which the major wild host grass species (selected in the preliminary survey above) were growing, were selected for these studies. The maize crop and wild host grasses were maintained by the farmer. Sampling was done once every three months starting July, 1997 to April, 1998, giving a total of 3 sampling occasions in each AEZ. A stratified random sampling procedure was used as explained below. Five sampling points were selected systematically along the terraces (field margins). The first sampling point was located at the edge of the first terrace, four other samples were taken at intervals of 5 - 10 metres apart depending on the length of each terrace. After the first terrace, sampling was done on next terrace (to the right), until the five sampling locations were selected. At each sampling point which was delineated using a 1m x 1m quadrant, 20 randomly selected tillers/plants were uprooted, thus giving a total of 100 samples per plot per sampling occasion. For each of the uprooted grass samples, the leaves and leaf sheaths were carefully examined for eggs and young larvae, while the stalks were dissected to locate older larvae and pupae. Larvae were divided into three size categories: small (1st and 2nd instars), medium (3rd and 4th instars) and large (5th and 6th instars), and placed individually in glass vials (2.5 cm diameter x 7.5 cm depth) containing artificial diet (Ochieng et al., 1985) and plugged with cotton wool. Pupae were placed individually in petri dishes (9 cm x 1.5 cm depth), lined at the bottom with moist filter paper. Egg masses (still attached to the oviposition substrate) were placed individually in petri dishes (9 cm x 1.5 cm depth), lined at the bottom with moist filter paper (3 cm x 3 cm). Humidity in the petri dishes containing the eggs and pupae was maintained by moistening the filter papers within, once every two days using a few drops of distilled water. All the stemborer samples were kept in the laboratory at ambient temperatures and reared to the adult stage. Emerged moths were examined to confirm the species identity. Parasitoids that emerged from the stemborers were identified. All voucher specimens were deposited at the International Centre of Insect Physiology and Ecology (ICIPE), Nairobi, Kenya for identification and classification. Data analysis. The percentage of each of the stemborer species and that of the various stemborer life stages collected was calculated for each of the wild grasses, in each of the zones. The number of each of the stemborer species was pooled for each of the wild grass species, and the overall percentage of each of the stemborer species and their life stages calculated for each of the wild grass species. Since parasitism was only observed in LM5, the percentage parasitism by various parasitoids was calculated for each of the stemborer species in this AEZ. For calculation of the the percentage parasitism, the susceptible stemborer life stages for each of the parasitoid species was used. For example, C. flavipes preferentially parasitizes the third to the sixth instar stemborer larvae (Smith et al, 1993). RESULTS Abundance and distribution of potential wild host grasses for stemborers. The abundance and distribution of the different potential wild host grasses of stemborers in the various AEZs is presented in Table 3. Within each AEZ, wild grass species whose abundance was over 70-% were: UM2 - Pennisetum purpureum SchumachPanicum maximum Jacq., UM3 - Pennisetum trachypyllum Pilg., P. purpureumand Panicum maximum, UM4 - P. purpureum, LM3 - P. purpureumand P. maximum, LM4 - P. purpureum and P. trachyphyllum, and LM5 - Sorghum versicola. These grasses were used in the subsequent on-farm study. However, although the area covered by Sorghum versicolor (LM5) Anders was less than 70%, this grass was selected since it was the most abundant wild host grass sampled in this site. Spatial distribution of stemborers in wild host grasses. Chilo partellus Swinhoe (Lepidoptera: Pyralidae) was the most prevalent stemborer that infested wild grasses in all the six zones considered together (Table 4). In LM5, where S. versicola was the major wild grass observed, the predominant stemborer recovered was C. partellus, which comprised 94.6% of the stemborers. No stemborers were recovered from any of the wild grasses in LM4. In LM3, P. maximum was mainly infested by Sesamia calamistis Hampson (Lepidoptera: Noctuidae), whereas only C. partellus was found in P. purpureum. On the contrary, C. partellus was the only stemborer that infested P. purpureum in UM4, and also P. purpureum and P. maximum in UM3 (Table 4). In UM2, C. partellus was again the main stemborer that infested P. purpureum and P. maximum, comprising 57.1% and 86.1% of the stemborers recovered from these grasses, respectively. All the wild grasses studied were vegetative throughout the season. Sorghum versicolor, P. maximum and P. purpureum were infested by stemborers throughout the season in the zones where they were recovered (Figs. 2 a - e and 3 a - c). In P. purpureum, the highest numbers of stemborers were recovered in January 1998 in LM3, UM3 and UM4. In P. maximum, the highest numbers of stemborers were recovered in January 1998 in LM3, whereas in UM2 and UM3, the highest numbers were recovered in July 1997. In S. versicolor, the highest numbers of stemborers were recovered in July 1997. Infestation of wild grasses. Chilo partellus was the dominant stemborer that infested P. purpureum, and the most commonly recovered stemborer life stage was the egg, followed by the medium-sized larvae (Table 4). Panicum maximum and S. versicolor were also most commonly attacked by C. partellus, with a majority of the stemborers recovered being in the medium- followed by the large-size larval category. No stemborers were recovered from Pennisetum trachyphyllum (Table 4). Parasitism of stemborers in wild grasses. Parasitoids were only recovered from LM5. In this single AEZ, parasitoids were recovered from C. partellus which infested S. versicola. Cotesia flavipes Cameron (Hymenoptera: Braconidae) was the most commonly recovered parasitoid (13.6%) (Table 5). The only small-size category larva recovered, was parasitized by Chelonus curvimaculatus (Cameron) (Hymenoptera: Braconidae), whereas parasitism by the other parasitoids was generally quite low (Table 5). No pupal or egg parasitoids were recovered. Parasitism by C. flavipes was observed from the first sampling in July 1997, and was highest in April 1998. DISCUSSION In this study, the major wild host grasses of stemborers were: P. purpureum, P. trachyphyllum, Panicum maximmum and Sorghum versicolor. A detailed account of 32 species of alternate wild host grasses of stemborers in the Coast, Nyanza and western Provinces of Kenya, has been given by Mathez (1972), Seshu Reddy (1983) and Khan et al. (1997). The range of alternate wild host grasses of stemborers, identified in this study was much narrower than that reported in the Coast, Nyanza and western parts of Kenya. It is likely that the comparative drier conditions in these semi-arid areas, limit the number of thick stemmed potential alternate host grasses that can survive in this region. In this study, P. purpureum was the most widely distributed wild host grass. This could be due to its widespread use as animal feed in various parts of eastern Kenya. Most farmers prefer it to the other fodder grasses due to its faster rate of development and higher dry matter content. In addition to being used as fodder for livestock, the wild grasses studied are also used for prevention of soil erosion and for stabilisation of terrace banks. This is the reason why these grasses are usually grown in strips along the edges of the bench terrace banks. In the present study, 18.2% and 14.3% of the stemborers at the pupal stage were recovered from P. purpureum and P. maximum, respectively. This contradicts earlier studies which showed that a very small percentage of stemborers develop to the pupal stage in these two wild grasses (Randriamananoro, 1996). Laboratory studies by Shanower et al. (1993) showed that only 1% and 0.3% of S. calamistis larvae developed to pupal stage, in P. purpureum and P. maximum, respectively. The foregoing suggests that there could be differences in suitability for stemborer development, among the different varieties of each specific grass species. It would therefore be useful in the future to evaluate the suitability of different varieties of wild host species for oviposition and development of specific stemborer species. This information can be used in the development of a habitat management programme for stemborers. Sorghum versicolor, was the most prefered wild host grass for stemborers, among the grasses studied. Mbapila (1996), reported that another species Sorghum arundinaceum (Desv.) Staf at the Kenyan Coast, was the most prefered alternate stemborer host grass species amongst those that were sampled (S. arundinaceum, P. maximum and P. purpureum). Field cage studies conducted by Randriamananoro (1996), showed that the wild sorghums, S. versicolor and S. arundinaceum were comparatively more attractive to B. fusca moths for oviposition, and were more suitable for settling by first instar larvae and larval development, than P. purpureum, P. trachyphyllum and the maize inbred line MP 704. It is possible that these two closely related wild sorghum species, have similar cues that attract oviposition by C. partellus, and also offer a suitable environment for the development of this stemborer. This suggests that wild sorghums have potential for use as trap crops, especially in fields planted with resistant maize varieties. There were comparatively more cases of the highest number of stemborers being recovered in January. This coincides with the non-cropping period, just after the short rains season (October - January). Towards the end of the short rains, maize enters the senescing stage and is thus less suitable for infestation by stemborers. It is conceivable that later generations of these stemborers migrate to the neighbouring actively growing alternate wild grasses, where they survive until the following long rains season (March - August), when some of them migrate back to the maize crop. In the present study, parasitoids were only recovered from the stemborers in S. versicolor, with C. flavipes being the most common parasitoid. The presence of C. flavipes in wild sorghum at Emali in LM5, was suprising since C. flavipes had not been released in the eastern Province of Kenya before the time of this recovery (from July, 1997 - April, 1998). In Kenya, C. flavipes was first released in the Kenyan Coast by the Commonwealth Institute of Biological Control (now known as CABI BioSciences) during the period 1968-1972; but did not establish (CIBC, 1968-1972). In 1993, C. flavipes was released again in the coastal area by ICIPE, and became successfully established (Overholt et al., 1997). The C. flavipes population in the eastern Province is likely to have originated from migrants arriving from the population established in the Coast Province since 1993. The area between the established population at the coast and the Eastern Province is mainly comprised of a non-cultivated, semi-arid landscape, most of which is part of Tsavo National Game Park. It is therefore possible that C. flavipes moved across this semi-arid stretch of land during seasonal windows of opportunity which occur during the rainy season when stemborers may infest thick-stemmed wild grasses. The gregarious endo parasitoid Cotesia sesamiae (Cameron) (Hymenoptera: Braconidae) was one of the indigenous parasitoids recovered. This study also showed that C. flavipes and C. sesamiae (that have been shown to successfully colonize C. partellus in maize), were able to survive successfully in S. versicolor. In laboratory experiments, C. flavipes and C. sesamiae were attracted to P. purpureum (Ngi-Song et al., 1995), and S. versicolor (Potting et al., 1995), suggesting that C. flavipes and C. sesamaie may be able to locate these wild grasses when actively growing maize is not available; such as after harvest, and during the off-season, when C. partellus are in aestivation in dry maize stalks (ICIPE, 1995; Randriamananoro, 1996.). Field cage studies by Mbapila (1996) showed that C. flavipes and C. sesamiae were unable to locate and parasitize aestivating larvae in dry maize stems. The availability of natural vegetation and weeds, which provide host larvae for adult parasitoids, have been reported to influence the dispersal of C. flavipes (Attique et al., 1980). In the present study, the exotic stemborer, C. partellus was the most widespread and most commonly recovered stemborer among the wild grasses infested by stemborers. This may well be due to the good colonization potential of C. partellus in new habitats (Overholt et al., 1994; Kfir, 1997). It is therefore likely that C. partellus adapts faster to the wild grasses and that it colonizes the suitable feeding niches in these habitats much earlier than the other stemborers, hence reducing the number of other stemborer species that successfully colonise these grasses. Chilo partellus, the dominant stemborer of maize in the semi-arid areas of eastern Kenya (Songa, 1999), is the co-evolved host for C. flavipes. Also, this study showed that C. partellus and C. flavipes can successfully survive in S. versicolor. The foregoing suggests that S.versicolor could be used in the management of stemborers in maize, by acting as a trap crop and also as a suitable refuge for C. flavipes, between cropping seasons. The integration of alternate wild grasses, such as S. versicolor into maize farming systems might assist in reducing the stemborer populations in maize farms in three ways. First, in areas where C. flavipes has been released and established, this parasitoid together with other indigenous species such as C. sesamiae, in the wild grasses, may reduce the initial available number of stemborers that may infest the maize crop at the beginning of the cropping season. Secondly, the wild grasses provide refuge and food for adult parasitoids during the non-cropping period of the year, enabling them to move back into the maize during the cropping season and parasitize the stemborers in the maize crop. Thirdly, the wild grass S. versicolor can act as a trap crop for the stemborers during the cropping season, thus reducing the number that infest the crops. Studies by Khan et al. (1997) in Nyanza province of Kenya, demonstrated that integration of Sudan grass Sorghum vulgare sudanense (Pers.) and molasses grass, Melinis minutiflora into maize cropping systems, significantly reduced stemborer infestation levels in the maize. They attributed this to the stimulo-deterrent diversionary effect of the two grass species, which involves repellance of insects from a harvestable crop with simultaneous attraction of the insects to a trap crop. This study revealed that in small scale farms in the semi-arid eastern Kenya, the wild grasses studied: S. versicolor, P. maximum, Pennisetum purpureum, P. trachyphyllum, had active vegetative growth throughout the year and were infested by stemborers during growth. Most farmers in this region practice mixed farming and rely on these grasses for animal feed (fodder) as well as free-range grazing throughout the year. These grasses are mainly grown along the edges of the terrace benches of the crop fields, where they serve the additional purpose of stabilising the terrace bank and preventing soil erosion. These considerations suggest that integration of the wild grasses into maize farming systems for stemborer management is consistent with the existing farming systems in this region, and therefore is very likely to be adopted by farmers in the semi-arid eastern Kenya. This study involved sampling of the wild grasses for one year, at intervals of three months. In order to develop a more sustainable habitat management system for stemborers in this region, there would be need for this study to be repeated, for about three more years, involving more intensive sampling. It would also be useful to test the suitability of sudan grass, Sorghum vulgare sudanense (Pers.), and molasses grass, Melinis minutiflora for use in the semi-arid areas of eastern Kenya. Studies in Nyanza Province by Khan et al. (1997) have demonstrated that these two grass species can facilitate the management of stemborers in maize cropping systems. ACKNOWLEDGMENTS We thank Dr. Susan Kimani and Mr. Tom Ondieki of The International Centre of Insect Physiology and Ecology (ICIPE) for identifying the parasitoids. Financial support for this work was provided by the Rockefeller Foundation. REFERENCES
©2002, African Crop Science Society The following images related to this document are available:Photo images[cs02018f1.jpg] [cs02018t2.jpg] [cs02018f3.jpg] [cs02018t3.jpg] [cs02018f2.jpg] [cs02018t4.jpg] [cs02018t1.jpg] [cs02018t5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}