 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 10. No. 3, 2002, pp. 195-201 ANALYSIS OF YAM YIELDS USING THE ADDITIVE MAIN EFFECTS AND MULTIPLICATIVE INTERACTION (AMMI) MODEL C. N. EGESI and R. ASIEDU1 Department of Crop Science & Technology, School of Agriculture and Agricultural
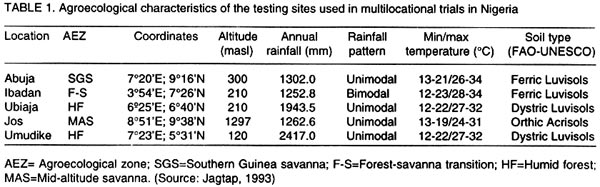
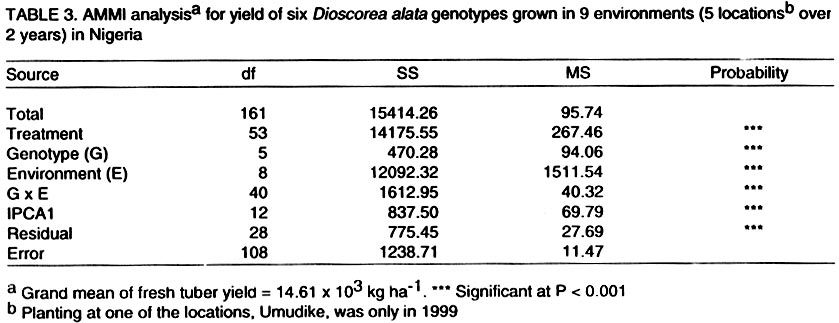
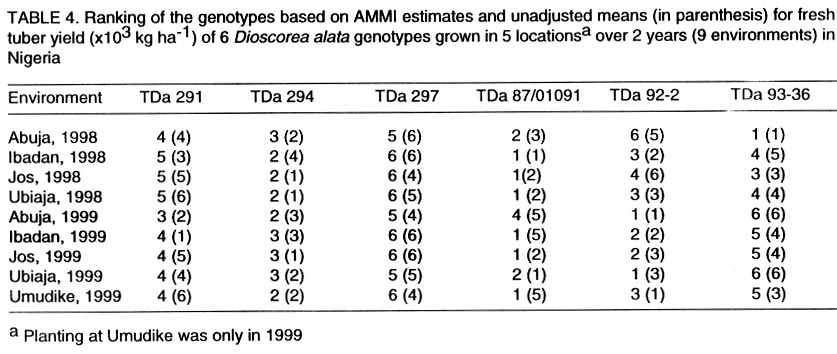
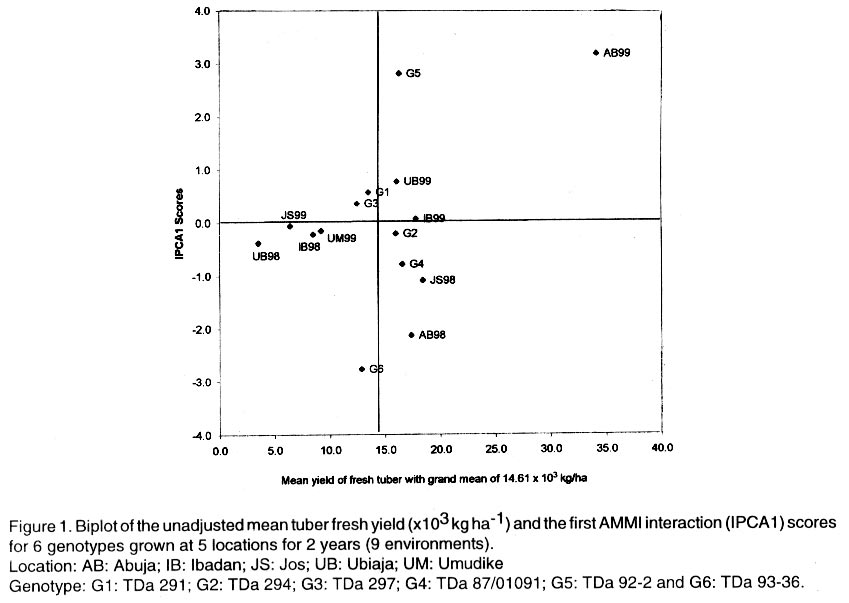
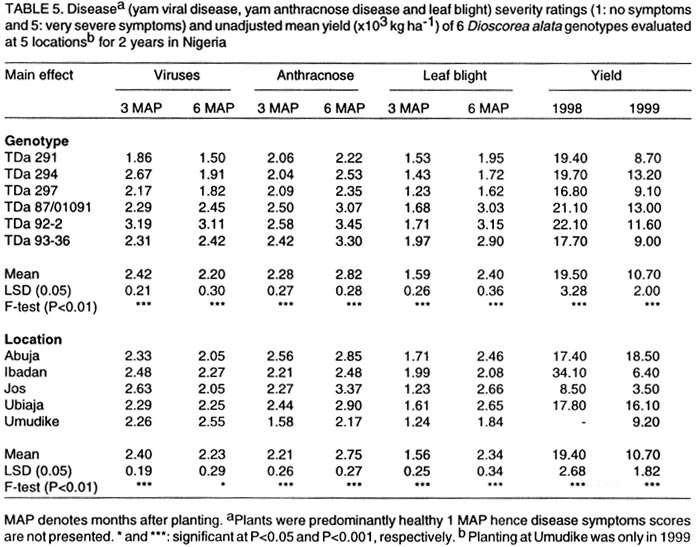
Technology, Federal University of Technology, PMB 1526, Owerri, Nigeria (Received 4 August, 2001; accepted 2 May, 2002) Code Number: cs02019 ABSTRACT The AMMI model combines regular analysis of variance for additive main effects with principal component analysis for multiplicative structure within the interaction. It also improves the accuracy of crop yield estimates and selects genotypes with highest yields. Despite knowledge of the model, potential yield estimates for yam cultivars as is the case for several other crops, are largely unknown. In this study, the AMMI model was used with the objective of assessing yam genotypes yield, selecting stable genotypes, and investigating G x E effects from trials conducted for two years at five locations in Nigeria. The effects of environments, genotypes and genotype-by-environment interaction (G x E) were highly significant (P<0.001). Within environments, AMMI1 estimates ranked genotypes differently from the unadjusted means, and in six out of nine cases AMMI1 estimates changed the top-yielding entry. The AMMI1 estimation also produced sharper and stratified rankings compared with the unadjusted means. The biplot showed four groupings of genotypes: TDa 93-36 generally low yielding and unstable; TDa 291 and TDa 297, low yielding and moderately stable; TDa 294 and TDa 87/01091 high yielding and stable; TDa 92-2 high yielding but unstable. Yam yields were relatively stable at Ibadan and Jos, while yields obtained at Abuja were unstable across both years. Superior yam selections with specific or broad adaptation were identified by AMMI. Key Words: Dioscorea alata, G x E, multilocational trials, yield stability RÉSUMÉ Le modèle AMMI combine l’analyse régulière de la variance pour les principaux effets additifs avec une analyse des composantes principales avec une structure d’intéraction multiplicative. Il améliore aussi l’estimation des rendements et la sélection des génotypes à haut rendement. En dépit de la connaissance du modèle, les estimations de rendements potentiels des variétés d’igname, comme c’est le cas de plusieures autres plantes, sont largement inconnues. Dans cette étude, le modèle AMMI était utilisé avec l’objectif d’estimer le rendement des génotypes d’igname, et d’examiner les effets de l’intéraction GXE dans les essais conduits pour deux ans dans cinq endroits différents. Les effets de l’environnement, génotypes et l’intéraction génotype et l’environnement était très significatifs (p<0.001). Dans les cinq endroits étudiés, les estimations AMMI ont classé les génotypes différement à partir des moyennes non ajustées, et dans six de neuf cas étudiés AMMI1 a modifié le classement des génotypes à haut rendement. L’estimation AMMI1 a aussi produit un classement stratifié pointu en comparaison avec les moyennes non ajustées. Quatre groupements des génotypes ont été obtenus par représentation graphique. La variété TDa 93-36 est généralement à rendement faible et instable, TDa291 et TDa297, à rendement faible et modérement stables ; TDa294 et TDa87/01091 à rendement élévé et stable ; et TDa92-2 à rendement élévé mais instable. Les rendements d’igname étaient relativement stables à Ibadan et Jos, alors que ceux obtenus à Abuja étaient instables pour les deux ans. Les sélections supérieures avec une adaptation spécifique et large étaient identifiées par l’AMMI1. Mots Clés: Dioscorea alata, GXE, essais multilocaux, stabilité du rendement INTRODUCTION One of the aims of yam (Dioscorea spp.) improvement programmes is to improve fresh tuber yields. Yams are grown in many agroecologies and it is important to assess adaptation and yield stability of promising genotypes across environments. Several methods have been proposed for statistical analysis of G x E and to predict the phenotypic response to changes in the environment (Eeuwijk, 1995). The success of identifying high yielding genotypes from yield trials depends on the effectiveness of the statistical analysis used to evaluate patterns in the data and estimated yields (Gauch and Furnas, 1991). The Additive Main Effects and Multiplicative Interaction (AMMI) model (Gauch, 1993) is more efficient in determining the most stable and high yielding genotypes in multi-environment trials compared to earlier procedures (Finlay and Wilkinson,1963; Eberhart and Russel,1966). The model uses the analysis of variance (ANOVA) approach to study the main effects of genotypes and environments, and a principal component analysis (PCA) for the residual multiplicative interaction between genotypes and environments. With the biplot facility from AMMI analysis, both genotypes and environments occur on the same scatter plot and inferences about their interactions can be made. The model focuses on the accuracy of estimates of yields of genotypes in multilocational trials. There is, however, limited application of the model. For instance, as is the case with other crops, yield stability for yams is largely unknown. This paper reports the use of AMMI analysis of yield data of six yam (D. alata) genotypes from two years of evaluation in five representative locations. The study was conducted with a view to establish patterns of responses of genotypes, environments and their interactions and to identify genotypes with broad or specific adaptation. MATERIALS AND METHODS The experiment was carried out in 1998 and 1999 at five locations in Nigeria. The locations included Abuja, Ibadan, Jos, Ubiaja and Umudike. At Umudike, the crop was grown only in 1999 giving a total of 9 environments. The agroecological characteristics of the locations, which represent major yam growing areas in Nigeria are shown in Table 1. Six genotypes from the working collection of IITA’s Yam Breeding Unit were used in this study, namely, TDa 291, TDa 294, TDa 297, TDa 87/01091, TDa 92-2 and TDa 93-36 (Table 2). At each location, the genotypes were grown in a randomised complete block design, with three replications under rainfed conditions. Planting was done at the beginning of the rains (May-June) at each location in both years with, 300 g setts. Each plot consisted of 40 plants in 8 rows (ridges of height 40 cm and 5 m in length) spaced I m apart. A plant spacing of 1 m x 1 m was used. No fertiliser or herbicides were applied during the course of the experiment. Hand weeding was done when necessary. Data on virus, yam anthracnose and leaf blight infections on foliage were estimated using a score scale of 1 to 5 (1 = no symptoms; 5 = very severe symptoms). The severity of disease symptoms was recorded at one, three and six months after planting (MAP). The tubers from each plot were harvested at 9 MAP and their fresh weights recorded. Data on disease scores were subjected to analysis of variance using SAS computer package (SAS, 1999). The AMMI statistical model (MATMODEL 2.0; Gauch, 1993) was then used to analyse the yield data. The AMMI model is: Yger = µ + ag + be + Sln ggn den + rge + Eger where Yger= yield of genotype g in environment e for replicate r; µ = grand mean; ag= mean deviation of the genotype g (genotype mean minus grand mean); and be = mean deviation of environmental mean; ln = the singular value for IPCA axis n; ggn = the genotype g eigenvector value for IPCA axis n; den= the environment e eigenvector value for IPCA axis n; rge = the residual; and Eger = the error. RESULTS AND DISCUSSION Genotypes (G), environments (E) and G x E effects were highly significant (P < 0.001) on yield and accounted for 3.32, 85.3 and 11.38% of the treatment sum of squares, respectively (Table 3). The postdictive success for AMMI using all data (all three replicates), indicated that the treatment sum of squares was partitioned into two components: 95.71% due to the model or pattern (including main effects for genotypes and the first interaction PCA) with 25 degrees of freedom; and 4.29% residual or random variation (noise) with 28 degrees of freedom. The model-validation procedure, therefore, identifies that the 4.29% of the treatment sum of squares as allocated to the residual term was not of predictive value. Within environments, AMMI1 frequently ranked genotypes differently from unadjusted means (Table 4). In six out of nine cases, AMMI1 estimation changed the top yielding entry. Thus, AMMI1 and unadjusted means selected the same winner in three out of nine environments, but selected different winners in the remaining six environments. With AMMI1 estimates, TDa 87/01091 ranked first in 6 environments against 2 given by the unadjusted means. For example, at Umudike, the unadjusted means ranked it fifth, while AMMI1 ranked it first. Thus, noise in the unadjusted means elevated some genotypes (such as TDa 92-2 and TDa 294) to higher positions. Previous yield trials on cassava (Manihot esculenta) (Dixon and Nukenine, 1997), soybean (Glycine max) (Gauch and Zobel, 1989) and maize (Zea mays) (Crossa et al., 1990) also showed that AMMI1 estimates differentially ranked top performing entries in over half the environments when compared with the unadjusted means. Therefore, AMMI1 estimation proved superior since ranking discrepancies between AMMI1 estimates and unadjusted means were due to random statistical variation. With the unadjusted means, genotype TDa 294 was generally in the top position (1-2) and also occurred three times in the middle rankings (3-4) but was never in the bottom rankings (5-6). With AMMI1 estimates, it was always in the top and middle positions. Similarly, TDa 87/01091 was 5 times in the top, once in the middle and thrice in the bottom. However, AMMI1 estimates generally placed this genotype mostly in the top (8 times) and once in the middle. Using the unadjusted means, TDa 297 was found to be 5 times in the middle and 4 times in the bottom, but occurred thrice in the middle and 6 times in the bottom with the AMMI1 estimation. The AMMI1 estimation had a profound effect in producing sharper and stratified ranking patterns and on this basis TDa 87/01091 would be considered more adapted to a wide range of environments than TDa 294 and the rest of the genotypes. Dixon and Nukenine (1997) and Crossa et al. (1991) obtained a similar stratification of cassava and wheat (Triticum aestivum) genotypes, respectively, using unadjusted means and AMMI1 estimates and noted that without AMMI1 estimation, noise in the data partially blurred adaptation patterns of genotypes to the extent that relatively well-adapted genotypes were grouped by chance in the middle and bottom positions. Conversely, a relatively poorly adapted variety sometimes occurred at the top for the same reason. The AMMI analysis provides a graphical representation or biplot (Fig. 1) to summarize information on the main effects and the first principal component scores of the interactions (IPCA1) of both genotypes and environments simultaneously (Kempton, 1984). Displacement along the abscissa reflected differences in main effects, whereas displacement along the ordinate exhibited differences in interaction effects. The AMMI1 expected yield for any genotype and environment combination can be calculated from Figure 1 following standard procedures suggested by Zobel et al. (1988). The additive AMMI0 which is part of the AMMI1 model is the sum of genotype and environment means minus the grand mean, while interaction part is the genotype PCA score times the environment PCA score. These two parts are added to produce the expected value of AMMI1 model. The biplot captured 94.5% of the treatment sum of squares. Genotypes or environments on the same parallel line, relative to the ordinate, have similar yields (Fig. 1), and a genotype or environment on the right side of the midpoint of this axis has higher yields than those on the left hand side. Consequently, the genotypes TDa 87/01091, TDa 92-2 and TDa 294 were generally high yielding, with TDa 87/01091 being the overall best. In contrast, TDa 297 and TDa 93-36 were generally low yielding genotypes. The Abuja location which was always on the right hand side of the midpoint of the main effect axis, seemed to be a favourable environment for the genotypes evaluated; Ibadan and Ubiaja were moderately favourable, while Jos and Umudike were less favourable environments. Genotypes with IPCA1 scores near zero had little interaction across environments and, vice versa for environments (Crossa et al., 1991). For such genotypes, overall mean ranking is a reliable measure. Genotype and environment combinations with IPCA1 scores of the same sign produced positive specific interaction effects, whereas combinations of opposite signs had negative specific interactions. Three genotypes TDa 87/01091, TDa 294 and TDa 291 had negligible interactions with the environments, indicating their broad adaptations with TDa 87/01091 being the most stable genotype across environments. Although, TDa 297 was fairly stable, it was not very productive. Genotypes, TDa 92-2 and TDa 93-36 were highly unstable and were generally poorly adapted to the environments. In 1999, G x E was generally low compared to 1998. Overall, Ibadan and Jos had lower IPCA1 scores than the other locations. Generally, Ubiaja and Umudike environments had moderate G x E interaction effects (average IPCA1 scores), while Abuja had the highest IPCA1 values and exhibited the most unstable yields. Thus, Abuja would be a good selection site for yam improvement when targeting a wide range of environments. Four groupings were evident from the biplot (Fig. 1): TDa 93-36 generally low yielding, and unstable (high negative IPCA1 score); and TDa 291 and TDa 297, were low yielding and moderately stable across environments (low positive IPCA1 scores); TDa 294 and TDa 87/01091 were high yielding, and stable (had low negative IPCA1 scores). Genotype TDa 92-2 was high yielding and very unstable across environments (high positive IPCA1 score). Stability in field performance of genotypes is influenced by prevailing biotic and abiotic stresses (Cock, 1985). Resistance to biotic stresses in addition to tolerance of common abiotic factors in the environments would ensure good varietal performance. In this study, the most stable genotypes (TDa 294 and TDa 87/01091) generally had moderate disease ratings (Table 5). On the other hand, TDa 92-2 (the most unstable genotype) and TDa 93-36 (a moderately unstable genotype) had the highest scores for yam anthracnose disease. Environments that exhibited stable genotype yields especially Ibadan, generally had lower anthracnose disease pressure compared with locations that exhibited unstable yields such as Abuja with higher temperature and relative humidity. The long growing period and diverse agroecologies in which yams are cultivated, expose cultivars to numerous biotic stresses with some being very devastating in cases of complex stress interactions. During the wet season, yam anthracnose disease and leaf blight can be very prevalent. They cause serious yield losses due to their destructive effects on the photosynthetic machinery of the plants. Yam improvement should be focused on multiple disease and pest resistance, which would guarantee stability of crop performance. Abuja had peculiar agroecological characteristics of high mean temperature and high relative humidity and exhibited unstable yam yields. Environments that showed moderate stability such as Ubiaja and Umudike are in the humid forest, have low altitude (210 m and 120 m, respectively), and are of the same soil type (Dystric Luvisols). Environments that exhibited stable yield: Ibadan (high yielding and Jos (low yielding), differed in their altitudes (210 m and 1,280 m, respectively) and mean temperature: factors which might have caused the differential stable yields. CONCLUSION The AMMI statistical model has been used to diagnose G x E interaction pattern of fresh tuber yield of yam. Genotypes TDa 87/01091 and TDa 294 were hardly affected by the G x E interaction effects and thus will perform well across a wide range of environments. Locations, such as Ibadan, that could be regarded as a good selection site for yam improvement due to stable yields observed were also identified. ACKNOWLEDGEMENTS The authors appreciate the assistance received from Dr. Umba-di-Umba of the Crop Improvement Division, IITA for transcribing the Abstract. We thank the staff of the Yam Breeding Unit, IITA for assistance in the field. Approved as IITA manuscript number IITA/01/JA/42. REFERENCES
©2002, African Crop Science Society The following images related to this document are available:Photo images[cs02019t4.jpg] [cs02019t2.jpg] [cs02019t1.jpg] [cs02019t5.jpg] [cs02019t3.jpg] [cs02019f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}