 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 10. No. 3, 2002, pp. 211-219 EFFET DE LA CONCENTRATION EN NACL SUR L’EMBRYOGENESE SOMATIQUE ET SUR LES CAPACITES DE REGENERATIONCHEZ LE BLE F. Oudija, M. Ismaili et M. Amsa Laboratoire de Physiologie Végétale et d’amélioration génétique des plantes, Département de Biologie, Université My Ismail, Faculté des Sciences, B.P. 4010, Meknès, Maroc (Received 27 July, 2001; accepted 21 March, 2002) Code Number: cs02021 RÉSUMÉ La réponse à la salinité a été étudiée chez deux variétés de blé dur Triticum durum (Karim et Sebou) et deux variétés de blé tendre Triticum aestivum (Sais et Merchouch) utilisées au Maroc. Le stress salin a été appliqué sur des cals de cinq semaines initiés sur un milieu de culture dépourvu de NaCl. La régénération a eu lieu dans des milieux de culture enrichis ou non en sel. Les résultats acquis montrent que les fortes concentrations en NaCl (10 et 15g l-1) ont perturbé les capacités embryogènes de tous les cultivars (P <0,0001). Ces concentrations ne sont pas létales. Cependant, on assiste à une diminution de la croissance relative et à une réduction de l’embryogenèse somatique avec l’augmentation de la concentration en sel dans le milieu de culture. A 15g l-1, la majorité des cals se nécrose et finit par mourir. Le blé tendre s’est montré plus tolérant que le blé dur (P<0,0001). Quant à la régénération, elle diminue avec l’augmentation de la concentration en sel dans le milieu de culture (P<0,0001). Les meilleurs résultats de régénération ont été obtenus dans le milieu dépourvu de sel et avec la concentration de 2,5 g l-1 de NaCl. A partir de 5 g l-1, les plantules déjà formées brunissent et le phénomène s’accentue avec l’augmentation de la concentration en sel du milieu. Le brunissement est total avec 15 g l-1 de NaCl. Mots Clés: Callogenèse, NaCl, salinité, stress, tolérance, Triticum aestivum, Triticum durum ABSTRACT Effect of salinity was investigated on four Moroccan wheat varieties: Karim, Sebou (durum wheat) and, Sais and Merchouch (soft wheat). Salt stress was applied to five week-old - calli which were cultivated in media deprived of salt. Regeneration was made in medium treated or not with salt. Results showed a decrease (P<0.0001) in embryogenesit ability at high salt concentrations (10 and 15g l-1) in all cultivars. These concentrations were not lethal. The higher salt stress was, the lower were relative growth and somatic embryogenesis. At 15g l-1, most calli showed necrosis and disappeared. Soft wheat tolerated more salt than durum wheat (P <0.0001). Regeneration was carried out in media supplemented or not with different amounts of salt. Regeneration decreased with these media salt concentration (P < 0.0001). The best regeneration results were obtained in media containing 0 or 2.5 g l-1 of salt. Browning started to show on the seedlings at 5g l-1 and was complete at 15g l-1. Key Words: Callogenesis, NaCl, salinity, stress, tolerance, Triticum aestivum, Triticum durum INTRODUCTION Le Maroc, dont une bonne partie des régions agricoles, se caractérise par un climat aride et semi – aride, est touché par la salinisation. Celle – ci prend de l’ampleur avec l’extension de l’agriculture irriguée. Une amélioration des germoplasmes cultivés au Maroc s’impose et demeure un moyen efficace de changement vers des variétés tolérantes pour disposer d’un matériel végétal susceptible d’assurer une production satisfaisante. L’emploi des techniques de sélection in vitro par isolement de lignées cellulaires tolérantes à la salinité constitue un moyen efficace pour imposer une pression de sélection à de nombreuses espèces végétales (Kochba et al., 1982). La culture de cellules sur des milieux strictement définis permet ainsi un traitement au sel précis et repérable ainsi qu’un contrôle stricte de leur environnement (Nabors et al., 1980, 1983). Des plantes entières ont été régénérées à partir de lignées cellulaires sélectionnées pour leur résistance à la salinité et la transmission de ce caractère de tolérance à leur descendance a été observée (Nabors et al., 1980; Collin et al., 1990). Parmi ces techniques, l’obtention d’une embryogenèse somatique in vitro par culture d’embryons immatures, permet la multiplication d’un génotype ainsi que la création de variants somaclonaux (Ozias-Akins et Vasil, 1982; Maddock et al., 1983). Les plantes régénérées à partir de ces derniers peuvent souvent présenter des caractères nouveaux, héritables et agronomiquement utiles et faire ainsi l’objet du développement de cultivars capables de tolérer la salinité. Le stade d’application du stress salin affecte le processus de sélection in vitro de variants tolérants. L’initiation des cals sur un milieu dépourvu de sel, suivie de plusieurs passages successifs sur des milieux sélectifs a permis l’obtention de lignées cellulaires n’ayant pas la capacité de retenir la tolérance à la salinité (Ben-Hayyim et Kochba, 1982; Hasegawa et al., 1980). Par contre Zair (1996), Oudija et Ismaili (2001), en travaillant sur le blé, ont pu constater que les cals initiés sur les milieux salins, présentent de meilleures performances embryogéniques qui conduiraient à la formation de lignées cellulaires tolérant mieux le chlorure de sodium. Ainsi, notre recherche a été consacrée à étudier l’effet du stress salin, imposé aux cultures de cals provenant d’embryons immatures de blé, dans le but de sélectionner des lignées cellulaires pouvant produire des embryons tolérants au chlorure de sodium et d’améliorer alors la tolérance des régénérants. MATERIEL ET METHODES Matériel végétal. Deux variétés commerciales de blé dur (Karim et Sebou) et deux variétés de blé tendre (Sais et Merchouch) ont été étudiées. Les graines ont été fournies par l’Institut National de Recherche Agronomique de Meknès (Maroc). La culture des plantes mères a été réalisée au champ sous des conditions naturelles. Les épis ont été prélevés 14 à 20 jours après l’anthèse. A ce stade, les embryons immatures ont une longueur de 1,5 à 2 mm et montrent une bonne morphogenèse in vitro (Bennici et al., 1988). La stérilisation des embryons immatures a été réalisée par passage dans un bain d’éthanol (95°C) pendant 30 mn suivi d’un bain d’hypochlorite de calcium (33%) pendant 15 mn. Les embryons ont été ensuite rincés 3 fois dans l’eau distillée stérile pendant 5 mn. Milieux de culture. Le milieu de culture de base (MS) se compose de sels minéraux et de vitamines selon Murashige et Skoog (1962) additionné de 30 g l-1 de saccharose, de 2 mg l-1 de 2,4 D (acide 2,4 dichlorophenoxyacétique) et de différentes concentrations en NaCl (0 - 2,5 - 5 - 10 et 15 g l-1 ). Ce milieu a été solidifié par 7 g l-1 d’agar. Le pH a été ajusté à 5,8 avant l’autoclavage. Les cals initiés sur le milieu MS pendant 5 semaines de culture ont été transférés sur des milieux salins contenant différentes concentrations en NaCl (0 - 2,5 - 5 - 10 et 15 g l-1 ). 30 cals furent utilisés pour chaque traitement. Le poids frais initial des cals a été déterminé en conditions stériles. Après 2 mois de culture dans les milieux salins, le poids frais final des cals a été déterminé de la même manière. Ainsi, on a pu déterminer les croissances relatives des cals selon la formule de Bhaskaram et al. (1983):
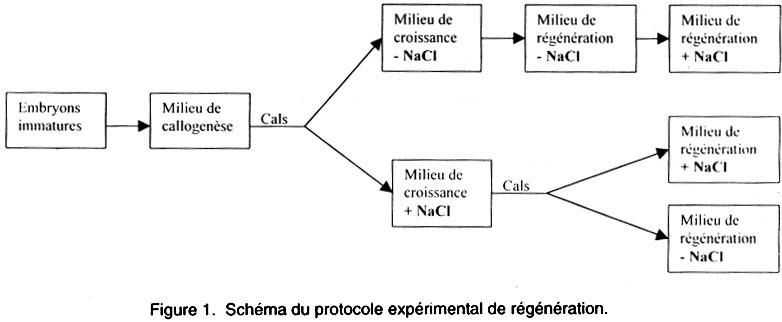
PF : poids frais; CR: croissance relative. Les cals induits dans le milieu MS sans sel, pendant 5 semaines de culture, puis transférés sur milieu de croissance avec NaCl, sont ensuite repiqués dans le milieu de régénération additionné ou non des concentrations suivantes en NaCl : 2,5 - 5 - 10 et 15 g l-1 selon le schéma suivant (Fig. 1). Conditions de culture. Les embryons immatures cultivés dans le milieu MS ont été soumis à une obscurité continue à une température de 25 ± 2°C le jour et 18 ± 2°C la nuit dans une chambre de culture pour l’induction de la callogenèse. Les cals ont été ensuite repiqués dans des bocaux de régénération additionnés ou non des différentes concentrations en sel. Ils ont été soumis à une photopériode de 16 heures d’éclairement. Evaluation des résultats. Après 5 semaines de culture, on a établi:
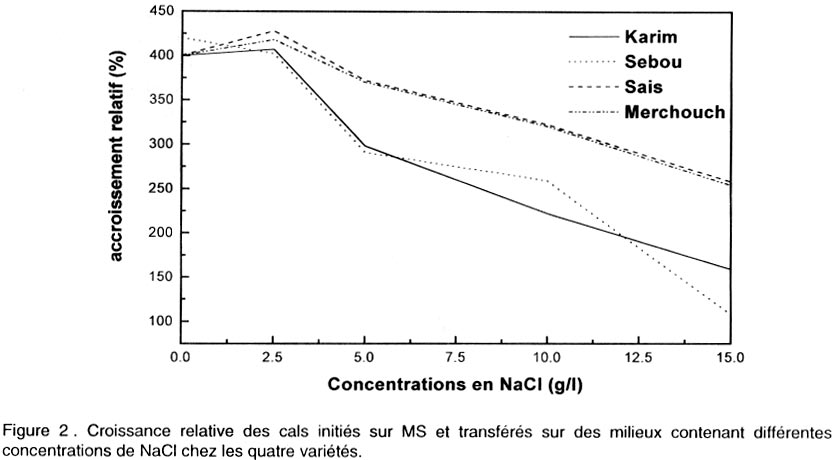
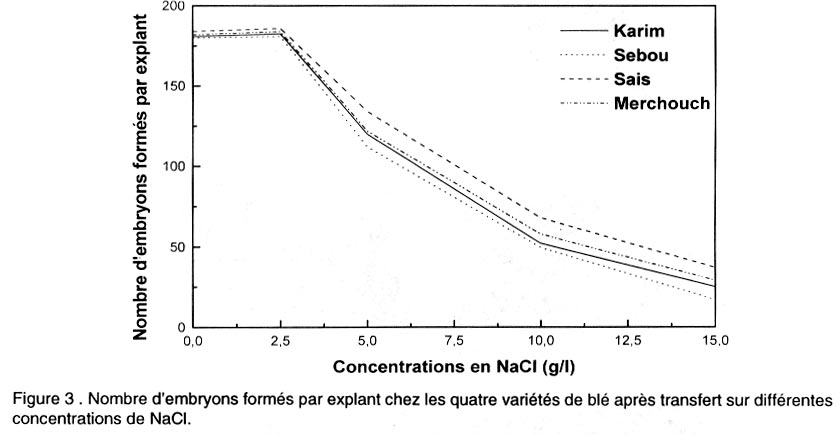
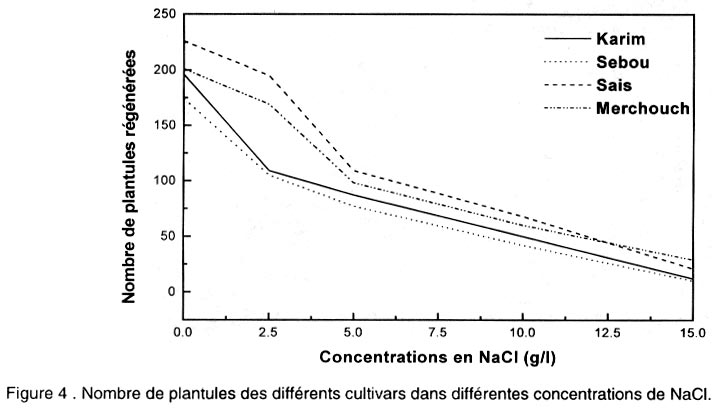
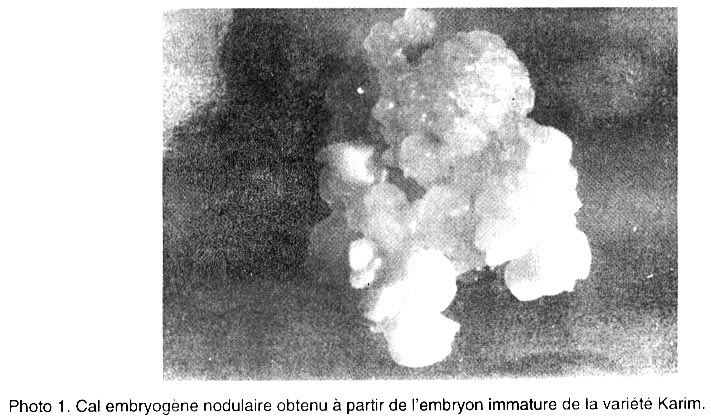
Le pourcentage de régénération est déterminé après 5 semaines de culture sur milieu de régénération par le nombre des cals ayant régénéré au moins une plantule par rapport au nombre total de cals mis en régénération. Analyse des données. L’analyse de la variance et la comparaison des moyennes ont été effectuées en utilisant le logiciel “SAS” (SAS, 1988). Une régression a aussi été appliquée sur les données afin de corréler certaines variables. RESULTATS Effet de NaCl sur la croissance des cals initiés sur le milieu MS Effet sur la croissance relative (CR). Les résultats de la Figure 2 montrent que le taux d’accroissement relatif des cals diminue progressivement et significativement avec l’augmentation de la concentration en NaCl (Tableau 1). Ainsi, la croissance des cals est ralentie par le stress salin. Pourtant, aucune différence n’a été observée entre les cultures témoins et celles du milieu contenant 2,5 g l-1 de NaCl. Le transfert des cals sur 2,5 g l-1 a légèrement stimulé l’embryogenèse chez la plupart des variétés. Par la suite, les compétences embryogènes des explants diminuent au fur et à mesure de l’augmentation du NaCl dans le milieu de culture. Le repiquage des cals sur 5 , 10 et 15 g l-1 de NaCl, est suivi d’une diminution significative du taux de croissance relative. Effet sur l’embryogenèse somatique. D’après la Figure 3, on constate que les nombres les plus élevés d’embryons somatiques formés par explant, ont été obtenus dans les milieux témoins et dans ceux contenant 2,5 g l-1 de NaCl. On note une légère stimulation de l’embryogenèse pour la dose 2,5 g l-1. Mais on n’enregistre pas de différence significative entre les deux milieux. Toutes les variétés réagissent ensuite à l’augmentation de la concentration en NaCl dans le milieu de culture, par une diminution progressive et significative des capacités embryogènes des explants (Tableau 1). Les nombres d’embryons par explant les plus faibles ont été obtenus avec les doses 10 et 15 g l-1 de NaCl. Dans ces milieux, on note la présence d’embryons de petite taille et dispersés dans la partie non embryogène du cal. Etude de la régénération à partir des cals initiés sur MS puis repiqués sur des milieux de croissance à différentes concentrations en NaCl Techniques 1 et 2. Chez toutes les variétés, les cals transplantés sur 0 et 2,5 g l-1 de NaCl régénèrent un grand nombre de plantules (Fig. 4). L’augmentation de la concentration en sel se traduit par une diminution progressive et significative du nombre de plantules formées pour toutes les variétés (Tableau 1). A 5 g/l, le nombre de plantules formées décroît fortement. Ce nombre est très réduit à 10 g l-1. Technique 3. Les embryons immatures des différents cultivars ont été mis en culture sur le milieu de callogenèse pendant 5 semaines. Les produits ont été ensuite transférés sur le milieu de régénération sans sel. Après un mois de culture, les plantes vertes régénérées ont été ensuite transférées sur le milieu de régénération contenant NaCl (0 - 2,5 - 5 - 10 et 15 g l-1). Au cours du premier mois de culture dans le milieu de régénération avec sel, un brunissement des plantules a apparu à partir de la concentration 5 g l-1. Ce brunissement est plus intense à partir de 10 g l-1 de NaCl, bien que les plantules vertes ont continué à y croître en produisant de nouvelles racines. Au cours de la deuxième subculture (deuxième mois de culture), le brunissement des plantules a été accentué. Après deux subcultures, aucune plante n’a survécu sur les milieux contenant 10 et 15 g l-1 de NaCl. Morphogenèse des cals. Les observations effectuées après 5 semaines de callogenèse à partir des explants immatures ont montré que la structure et la morphologie des cals ont été affectées par la concentration en sel. L’aspect des cals obtenus a varié selon la dose de NaCl utilisée. Les cals repiqués sur MS additionné de 2,5 g l-1 de NaCl présentent la même structure morphologique que ceux obtenus dans le milieu témoin. Ils ont été bien développés, embryogènes et constitués de deux parties (voir photo 1):
Dans le milieu MS supplémenté de 5 g l-1, les cals ont été identiques à ceux décrits précédemment mais avec prédominance de la partie embryogène. On a constaté aussi un début de brunissement des cals chez la variété Merchouch. Après 6 semaines de culture dans le milieu MS + 10 g l-1 de NaCl, on a observé un changement de l’aspect des cals. Chez certains cultivars, le brunissement a été prononcé et la plus grande partie du cal a été non embryogène; c’ était le cas de Sais et Merchouch. Les autres variétés (Karim et Sebou) ont semblé bien tolérer la dose 10 g l-1 de NaCl et ont présenté une partie embryogène dominante. Dans MS + 15 g l-1 de sel, la plupart des cals se nécrosent. Ils deviennent friables, mous et bruns (photo 2). Chez les variétés résistantes, les cals ont conservé une petite partie embryogène comme dans le cas de la variété Sais (photo 3). DISCUSSION ET CONCLUSION Les résultats de nombreux travaux ont montré que les techniques de culture in vitro sont les plus adéquates pour la sélection de lignées cellulaires tolérantes à la salinité (Zenk, 1974; Dix, 1980; Kochba et al., 1982). Les variants sélectionnés à ce jour pour la tolérance à la salinité ont été obtenus par des méthodes de sélection fondées sur la mise en œuvre d’une pression sélective inhibant la croissance et tuant les cellules sensibles (Maliga, 1984). La pression sélective est obtenue en supplémentant le milieu de culture avec différentes concentrations en sel. Les cellules tolérantes, identifiées par leur croissance ou leur survie en présence du sel, sont transférées sur un milieu de régénération additionné ou non en sel. L’application de l’agent stressant peut avoir lieu à différentes phases de croissance des cals. En effet, l’initiation des cals sur un milieu ne contenant pas de sel, suivie de plusieurs passages successifs sur des milieux sélectifs a été étudiée par plusieurs chercheurs. Ben-Hayyim et Kochba (1982) ont pu obtenir une lignée (R - 10) tolérante à NaCl après irradiation des cellules par des rayons (12 k rad) suivi de transferts successifs sur un milieu contenant 0,2 M de NaCl. De même, Hasegawa et al. (1980 ) ont isolé une lignée cellulaire (S-10) dont le poids de matière fraîche et sèche obtenu sur un milieu contenant 10 g l-1 de NaCl est de 60 à 100 % respectivement par rapport aux masses obtenues par la même souche sur le milieu témoin. Cependant, dans les deux cas (R - 10 et S - 10), après transfert sur des milieux dépourvus de sel, les cellules de ces lignées ont perdu leur tolérance à NaCl et ont répondu comme les cellules non sélectionnées. D’autres chercheurs ont pu sélectionner des variants ayant la capacité de retenir la tolérance à la salinité. C’est le cas de Beloualy et Bouharmont (1992) qui ont sélectionné des lignées cellulaires de Citrus (Poncirus trifoliata) sur des milieux de culture contenant 5 et 10 g l-1 de NaCl. Ces lignées cellulaires ont montré une croissance normale pour les concentrations utilisées dans la sélection. Les plantes régénérées à partir de ces lignées ont aussi montré une amélioration de la croissance et de la tolérance à la salinité. D’autres chercheurs se sont intéressés à l’application du stress salin pendant la phase d’initiation des cals. C’est le cas de Zair (1996) en travaillant sur le blé. Il a pu constater que ces cals présentent de meilleures performances embryogéniques. Ceci pourrait s’expliquer par le fait que l’application du sel en phase d’initiation des cals pourrait provoquer des mutations qui conduiraient à la formation de lignées cellulaires tolérant mieux le chlorure de sodium. Ces observations confirment les résultats d’Abrigo et al. (1985). La présence de l’agent stressant dans le milieu de régénération peut aussi augmenter la probabilité de régénérer des plantes tolérantes (Nabors et al., 1980). Le stress salin peut être également exercé graduellement, par palier, en augmentant la concentration du sel au cours des repiquages successifs des colonies survivantes (Meredith, 1984). La pression sélective graduelle peut cependant favoriser des mécanismes d’adaptation physiologiques (McHughen et Swartz,1984; McHughen, 1987) qui ne manifestent plus leurs effets au niveau des plantes régénérées, ce qui n’est pas souhaitable dans un programme de sélection. Pour les blés, un brunissement apparaît dans le milieu de régénération contenant les fortes concentrations en NaCl. Le brunissement résulte d’une mise en contact des systèmes oxydatifs, essentiellement cytoplasmiques, et de leurs substrats, plutôt vacuolaires (Mayer et Harel, 1981). Ce phénomène implique une décompartimentation cellulaire consécutive à des traumatismes mécaniques (coupes, blessures,…), à la sénescence des tissus ou à une dégradation accidentelle des membranes par des formes toxiques d’oxygène. Ce brunissement est lié essentiellement aux systèmes enzymatiques classiquement impliqués dans l’oxydation des composés phénoliques. En effet, les recherches menées dans ce sens sur Hevea brasiliensis en culture in vitro, ont montré que le brunissement peut s’expliquer globalement par une augmentation des activités peroxydases (néfaste) et une diminution de l’activité catalase (protectrice) (Housti et al., 1991). En guise de conclusion, nos résultats montrent à la fois une réduction de la croissance des cals, de leur compétence embryogène et des capacités de régénération au delà de la concentration de 2,5 g l-1 de NaCl. Cette réduction s’accentue avec l’augmentation du stress salin dans le milieu de culture. D’autre part, aucune variété n’a été affectée par la dose 2,5 g l-1, bien au contraire, la croissance des cals est stimulée chez toutes les variétés. De plus, cette concentration est insuffisante pour distinguer les variétés résistantes des autres. Les concentrations de 5 et 10 g l-1 de NaCl ne sont létales ni pour les variétés de blé dur ni pour celles du blé tendre. Cependant, on note une réduction de la croissance par rapport au témoin qui s’accentue encore avec 10 et 15 g l-1 de NaCl. Le blé tendre s’est montré plus tolérant que le blé dur. Dans le milieu de régénération supplémenté de fortes concentrations en chlorure de sodium, un brunissement apparaît chez toutes les variétés de blé étudiées. REFERENCES
©2002, African Crop Science Society The following images related to this document are available:Photo images[cs02021f1.jpg] [cs02021t1.jpg] [cs02021f2.jpg] [cs02021f3.jpg] [cs02021p1.jpg] [cs02021f4.jpg] [cs02021p2.jpg] [cs02021p3.jpg] |
| |||||||||


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}