 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 10. No. 4, 2002, pp. 281-310 THE PERSPECTIVE OF SWEETPOTATO CHLOROTIC STUNT VIRUS IN SWEETPOTATO PRODUCTION IN AFRICA: A REVIEW R. W. GIBSON and V. ARITUA 1 Natural Resources Institute, University of Greenwich, Central Avenue,
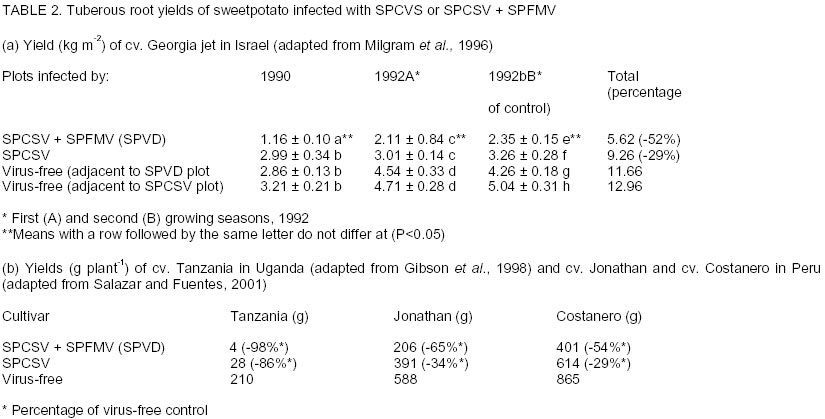
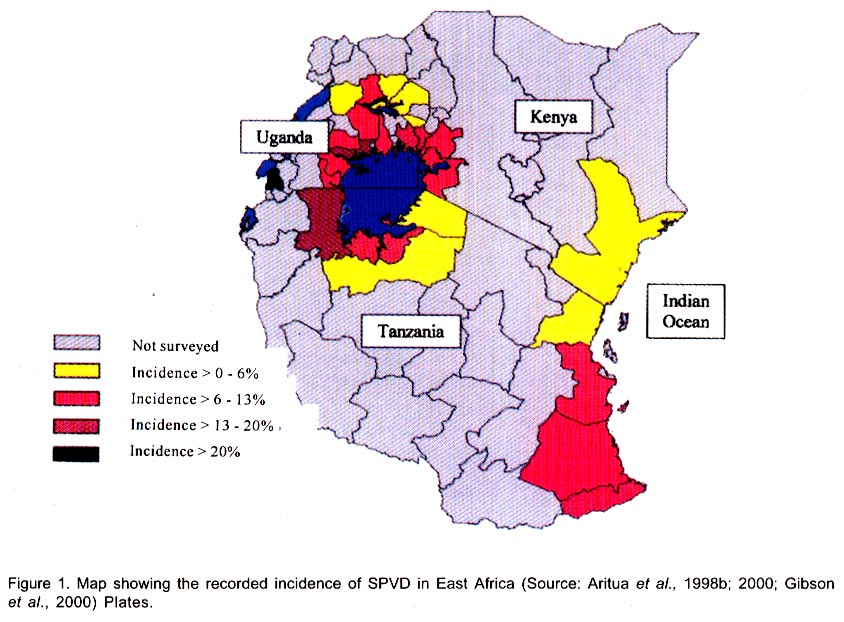
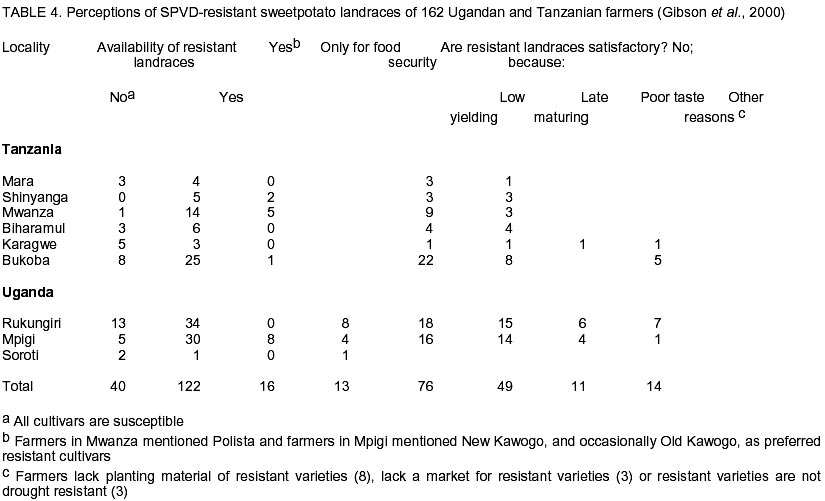
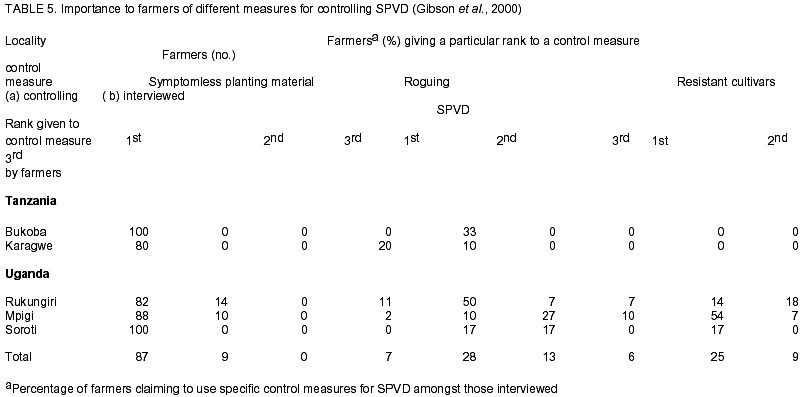
Chatham Maritime, Kent, ME4 4TB, UK. (Received 5 August 2002; accepted 8 October, 2002) Code Number: cs02028 ABSTRACT Sweetpotato chlorotic stunt virus (SPCSV) (Crinivirus: Closteroviridae) occurs in the main tropical regions of the World and is probably the most damaging pathogen of sweetpotato (Ipomea batatas). However, until recently, little research had been done on it. The methods of identification were inadequate and this led to synonymy. The virus is transmitted by the whitefly species, Bemisia tabaci and Trialeurodes abutilonea, in a semi-persistent fashion. At least two serotypes occur, one, first described from West Africa (SPCSV WA ), and the other first described from East Africa (SPCSV EA ). Both serotypes have also been found in the Americas. Nucleotide sequencing has facilitated the process of distinguishing strains. The SPCSV may have originated along with sweetpotato in the Americas, but is perhaps more likely to have been a ‘new encounter’ for sweetpotato when it was introduced to Africa and elsewhere in the ‘Old World’. It infects few plant species other than Ipomoea spp. The virions comprise long flexuous particles and the genome is RNA and bipartite. Geographically, isolated strains of SPCSV have been distinguished using serological- and nucleic acid-based methods. The virus synergises Sweetpotato feathery mottle virus (SPFMV) (Potyvirus: Potyviridae), leading to increased titres of this virus and the development of the severe disease sweetpotato virus disease (SPVD) in dually infected sweetpotato plants. Plants affected by SPVD following artificial or natural infection with SPCSV plus SPFMV have generally yielded c.60 –90% less than uninfected or unaffected controls in field trials; plants infected with SPCSV alone generally yielded c.30 – 80% less. However, in crops, compensatory growth by unaffected neighbouring sweetpotato plants probably results in direct effects on overall crop yields being small, and the major effect of SPCSV in constraining the yields of sweetpotato is perhaps through preventing the cultivation of high yielding but SPVD-susceptible sweetpotato cultivars. Breeding resistant high-yielding varieties has been the main means of avoiding the deleterious effects of SPVD. Although sweetpotato cultivars which have field resistance to SPCSV have been identified, no gene conferring immunity to SPCSV has been identified within sweetpotato or its close relatives. Cultivars non-indigenous to Africa seem particularly susceptible. Extreme resistance has been identified in certain wild Ipomoea spp. Recent epidemiological studies indicate that most spread of SPCSV is short distance, leading to interest in the use of phytosanitation measures to grow high-yielding but somewhat SPVD-susceptible cultivars. This paper comprises a review of research work done on SPCSV world wide dating from 1939 to-date. Key Words : Africa, Bemisia tabaci, Ipomoea batatas, Sweetpotato feathery mottle virus RÉSUMÉ Le virus d’arrêt de croissance de la patate douce chlorotique (SPCSV) (Crinivirus: closteroviridae) apparaît dans les principales régions tropicales du monde et est probablement le pathogène le plus préjudiciable de la patate douce (Ipomea batatas). Cependant, jusque récemment, peu de recherches avaient été faites sur lui. Les méthodes d’identification étaient inadéquates et ceci conduit a la synonymie. Le virus est transmis par les espèces de mouche blanche, Bemisia tabaci et Trialeurodes abutilonea, dans un mode semi persistant. Au moins deux sérotypes ont lieu, un, premièrement décrit en Afrique de l’ouest (SPCSV WA ) et l’autre en Afrique de l’est (SPCSV EA ). Les deux sérotypes ont aussi été trouvés en Amérique. La nucléotide mise en séquence a aussi distingué les souches. Le SPCSV peut avoir tiré son origine avec la patate douce en Amérique, mais est peut être plus vraisemblablement un nouveau rencontré pour la patate douce quand il était introduit en Afrique et ailleurs dans le vieux monde. Il infecte peu d’espèces de plantes autre que l’Ipomea spp. Les virions comprennent des longues particules de flexuous et le génome est le RNA et le bipartite. Géographiquement, les souches isolées de SPCSV ont été distinguées en utilisant les méthodes basées sur l’acide serologique et nucléique. Les virus synerge, le virus plumeux de taches de patate douce (SPFMV) (Potyvirus: Potiviridae) menant à des titres croissant de ce virus et le développement de la sévère maladie de virus de la patate douce (SPVD) en plantes de patate douce doublement infectées. Les plantes infectées par SPVD artificiellement ou naturellement avec SPCSV en plus du SPFMV ont généralement produit 60-90% moins que celles non infectées ou non affectées en champ d’essais. Les plantes infectées avec le SPCSV seul, généralement ont produit 30-80% moins. Cependant, en plantes, la croissance compensatoire par les plantes de patate douce voisines non affectées résultent dans les effets directs sur les rendements totaux des plantes étant moindres, et l’effet majeur de SPCSV en contraignant les rendements de patate douce est peut être a travers la prevention de cultures de haut rendement mais le SPVD prédispose la patate douce aux cultivars. La production des variétés résistantes de rendement élévé a été le moyen principal d’éviter les effets nuisibles de SPVD. Bien que les cultivars de patate douce qui ont un champ de résistance au SPCSV ont été identifiés, aucun géne conférrant l’immunité au SPCSV a été identifié dans les patates douces ou leurs proches. Les cultivars non indigènes à l’Afrique semblent particulièrement prédisposés. Une extrême résistance a été identifiée dans certains Ipomea spp. sauvages. Des récentes études épidémiologiques indiquent que le plus repandu de SPCSV est à courte distance, menant à l’intérêt dans l’usage des mesures phytosanitaires pour planter des variétés à rendement élévé et quelque peu prédispose au SPVD. Cet article comprend une revue de travaux de recherche fait sur le SPCSV dans le monde entier de 1939 a nos jours. Mots Clés : Afrique, Bemisia tabaci, Ipomea batatas, Virus plumeux de tache de patate douce INTRODUCTION Sweetpotato (Ipomoea batatas (L.) Lamarck), the third most important tuberous root crop in the world, is grown predominantly in tropical and semi-tropical developing countries. China has the largest production followed by Uganda (FAO, 1999). Despite this, Africa produces only about 6% of the world crop, but almost all the crop is consumed directly by humans hence the crop has a relatively large nutritional impact. In Africa, the tuberous root is eaten mostly after boiling or steaming the freshly harvested tubers, although in areas with prolonged drought, it may also be preserved as dried chips (Bashaasha et al., 1995; Kapinga et al., 1995). In some African cultures, leafy shoots are eaten. Sweetpotato is especially important in the countries surrounding Lake Victoria; per capita consumption is greatest in Rwanda(FAO, 1999). Production is increasing throughout much of Africa in response to increasing human populations and market liberalisation, removing such factors as subsidies to fertilisers and seed or guaranteed prices which previously benefited high input crops. Pests and diseases are also adversely affecting the production of alternative staples, notably cassava mosaic geminiviruses affecting cassava; black sigatoka, banana (Musa spp.) weevil and nematodes affecting bananas; and grey leaf spot affecting maize. Sweetpotato production is being encouraged by national governments and by non-government organisations (NGOs) for food security as the crop yields quickly following drought and, in additon, yields relatively well even in nutrient-depleted soils. Sweetpotato is grown primarily by smallholder farmers, often in localities vulnerable to drought and on impoverished soils (Bashaasha et al., 1995; Kapinga et al., 1995). The ability of the crop to establish and yield quickly provides both ground cover and an early source of food following the onset of rains. The identification of orange-fleshed, high vitamin A varieties (Anonymous, 2000) further adds to the benefits of the crop in countries where high proportions of the population, particularly women and children, suffer from vitamin A deficiency even to the extent of causing blindness. Sweetpotato is mostly grown and eaten by the poorest among African communities. It is grown especially by women, for daily family consumption and for sale. Therefore, increased production contributes directly to poverty reduction in much of Africa (Tardif-Douglin, 1991; Bashaasha et al., 1995; Kapinga et al., 1995; FAO, 1999; Hakiza et al., 2000; Scott et al., 2000). Sweetpotato virus disease (SPVD) is the main disease of this crop in Africa (Geddes, 1990) and elsewhere (Carey et al., 1999). Despite the apparent broad meaning of the name, SPVD has become associated closely with symptoms caused by a combination of two viruses: Sweetpotato chlorotic stunt virus (SPCSV) (Crinivirus; Closteroviridae) and Sweetpotato feathery mottle virus (SPFMV) (Potyvirus; Potyviridae) (Gibson et al., 1998a). The symptoms include an often severe stunting of the plant and stunting, distortion and either a chlorotic mottle or vein clearing of the leaves. The tuberous root yield of affected plants is also small. The significance of the aphid-borne SPFMV in sweetpotato production in Africa has recently been reviewed (Karyeija et al., 1998a); and in this paper we arguably present a review of the most economically important whitefly-borne component, SPCSV, again focusing on Africa. This paper provides a review of literature on this virus and its impact on sweetpotato production world wide ranging from 1939 to-date. HISTORICAL REPORTS OF SPCSV AND SPVD The first published description of SPVD was from Uganda (Hansford, 1944). Affected sweetpotato plants were described as having severe stunting with a chlorotic mottle or mosaic affecting the leaves which were often also stunted and bronzed; almost complete loss of tuberous root yield was also reported. Large numbers of Bemisia spp. whiteflies were present on the crop. This was apparently the first report of this insect on sweetpotato in Uganda and of a possible linkage between whiteflies and SPVD. The SPVD-affected plants were common and widespread. Reports of SPVD from Ruanda-Urundi (now Rwanda and Burundi), western Tanganyika (probably referring to what is now Kagera district in Tanzania) and Congo Belge (now Democratic Republic of Congo) were also mentioned (Sheffield, 1953). In Congo Belge, SPVD caused sweetpotato to be abandoned as a major food crop in the Kilo mines at Ituri (west of Lake Albert), the annual crop production being reduced from 30,000 kg to 4,000 kg in just a few years. This disaster, which occurred around 1939, provides probably the first written record of SPVD in Africa (reported by Stayaert; see Sheffield, 1953). Furthermore, “During this same period, farmers near Mulungu-Tshibinda [also in D. R. Congo] on the western shores of Lake Kivu began to complain of degeneration in their sweetpotato” yields. Sheffield (1953) personally observed SPVD in Uganda, Belgian Congo, Ruanda-Urundi, Kenya and Tanganyika and suggested from its widespread distribution that it had probably been in Africa for a considerable time. Since then, SPVD has been reported in Ghana (Clerk, 1960), Nigeria (Schaefers & Terry, 1976), Cameroon (Ngeve & Boukamp, 1991), Madagascar, Zambia (Gibson et al., 1998a), Togo, Liberia, Sierra Leone, São Tomé, Ivory Coast (Thottappilly & Rossel, 1988), Benin and Gabon (Lenné, 1991). During the 1950s - 1960s, whitefly-borne viruses were isolated from sweetpotato in different parts of the world and, in the absence of specific assays, given different names. This approach continued, but more slowly, into the 1990s after which diagnostic assays based on viral RNA or coat protein, allowed relationships to be assessed more accurately. Isolates of these early named viruses seem not to have been retained, so whether they are synonymous with SPCSV remains unclear. In East Africa, Sheffield (1957, 1958) detected two viruses in sweetpotato affected by SPVD. Both viruses (called virus A and virus B) were isolated from diseased sweetpotato samples originating from Kenya, Tanganyika Territory (now part of Tanzania) and Uganda. Virus A was aphid-borne and is now considered to have been SPFMV, while, virus B was transmitted by the whitefly B. tabaci (Gennadius) (Karyeija et al., 1998a). Cohen et al. (1992) suggested that virus B was SPCSV largely because SPCSV is the ubiquitous whitefly-borne component virus of SPVD (Gibson et al., 1998b). However, virus B was transmitted by sap, and to diverse hosts including Gomphrena globosa, Mina lobata, Datura ferox, Lycopersicon esculentum, Petunia sp. and Physalis peruvianum (Sheffield, 1958). Neither sap transmission, hosts other than Ipomoea spp., Amaranthus spp. and Nicotiana spp., nor the symptoms described (mottling, vein clearing, necrotic lesions, etc.) are characteristic of SPCSV (Cohen et al., 1992). However, they are characteristic of the whitefly-borne virus, Sweetpotato mild mottle virus (SPMMV) (Ipomovirus; Potyviridae), which also occurs in East Africa and which may also be Sheffield’s virus B (Hollings and Stone, 1976). The SPMMV can occur in complex mixtures with SPCSV and SPFMV in diseased sweetpotato plants (Gibson et al., 1998b). Thus, virus B may also have been a complex of SPCSV and SPMMV, which would account for the wide host range of virus B and the diverse isolates reported (Sheffield, 1958). A probable virus which is whitefly-borne and causes a leaf curl has also been reported in Nigeria but the symptoms seem distinct from those of either SPVD or SPCSV (Thottappilly and Rossel, 1988). In the U.S.A., sweetpotato mosaic transmitted by B. tabaci affected sweetpotato in south Georgia (Girardeau, 1958; Girardeau and Ratcliffe, 1960). Yellow dwarf, a virus transmitted by the whitefly Trialeurodes abutilonea (Hald.) and, which caused “sweetpotato feathery mottle virus complex” in combination with SPFMV, has also been reported to infect sweetpotato in Maryland (Hildebrand, 1960). Either one or both whitefly-borne viruses may have been synonymous with SPCSV as SPCSV is now known to occur in the U.S.A (Pio-Ribeiro et al., 1996) and to be transmitted by B. tabaci and T. abutilonea (Valverde, 1999; Sim et al., 2000). However, the geminivirus Sweetpotato leaf curl virus (SPLCV), also occurs in sweetpotato in U.S.A., is transmitted by B. tabaci (Lotrakul et al., 1998) and so may instead have been involved in the aetiology of sweetpotato mosaic. Confusingly, a whitefly-borne sweetpotato virus from Taiwan has also been called yellow dwarf (Green, 1987), though its relationship to the yellow dwarf of Hildebrand (1960) seems unclear. In Israel, sweetpotato vein clearing virus (SPVCV) disease has a whitefly-borne component (Loebenstein and Harpaz, 1960) which may be synonymous with sweetpotato sunken vein virus (SPSVV) (Cohen et al., 1992), itself synonymous with SPCSV (Gibson et al., 1998b), although the name SPSVV is still used occasionally (Cohen et al., 2001). The first unambiguous isolation of SPCSV was in Nigeria (Schaefers and Terry, 1976), using B. tabaci to transmit it from sweetpotato with SPVD to I. setosa. They named the virus Sweetpotato chlorotic stunt virus from the symptoms it induced in I. setosa and preliminarily identified it as a closterovirus. They similarly used aphids to isolate a virus now known to be SPFMV from SPVD-affected sweetpotato. Graft-transmitting both SPFMV and SPCSV back to single virus-free sweetpotato plants induced SPVD. Isolates of Nigerian SPCSV have been maintained and riboprobes developed to provide a consistent detection method. It has been confirmed to be a closterovirus (hence, SPCSV has also been called SPVD-associated closterovirus (Winter et al., 1992a, b). The name Sweetpotato chlorotic stunt virus has since been accepted by the Closterovirus Study Group of the International Committee for the Taxonomy of Viruses. That SPCSV is consistently associated with SPVD and that dual infection with SPCSV and SPFMV are required to induce SPVD symptoms has since been confirmed in East Africa (Gibson et al., 1998b). Two serologically-distinguishable and geographically-separated strains of SPCSV have been identified in Africa. In this review, the strain found in West Africa is referred to as SPCSVWA because it was first isolated from West Africa (Nigeria) (Schaefers and Terry, 1976). Similarly, the strain found in East and Southern Africa (Gibson et al., 1998 a & b) is referred to as SPCSVEA because it was first isolated from East Africa (Kenya) (Hoyer et al., 1996). The name sweetpotato chlorotic stunt has also been used inadvertently for a potyvirus from the Caribbean (Brown et al., 1988; Lenné, 1991). This virus has also been identified serologically in East Africa (Wambugu et al., 1991), though not confirmed. Antiserum against this virus failed to detect it in sweetpotato in Zimbabwe (Chavi et al., 1997). GEOGRAPHICAL DISTRIBUTION OF SPCSV As mentioned above, the first records of SPCSV and/or SPVD are from Africa. The disease was reported in 1939 in eastern D. R. Congo, in almost the centre of Africa, and had been observed throughout at least East Africa by the 1940s (Sheffield, 1957). It was also first named from West African isolates (Schaefers and Terry, 1976) and both SPCSVEA and SPCSVWA were first reported in sub-Saharan Africa. However, SPVD was probably reported, albeit more rarely and under other names, in other continents during the 1950’s (see Historical Reports of SPCSV and SPVD). More recently, SPCSV has been identified in Egypt (Carey et al., 1999), Israel (Cohen et al., 1992), U. S. A. (Pio-Ribeiro et al., 1996), Brazil (Pio-Ribiero et al., 1994), Argentina, Peru on both the Pacific and Atlantic sides of the Andes, west, south and east Java and Indonesia (Carey et al., 1999) and Taiwan (Winter et al., 1997). The SPCSVWA, as well as being found in West Africa, has also been found in Israel, U.S.A. (Hoyer et al., 1996) and Argentina. Similarly, SPCSVEA, which occurs in East and southern Africa from Uganda and Kenya (Gibson et al., 1998b) Zambia and Madagascar (Gibson et al., 1998a), also occurs in Peru in South America (Carey et al., 1999). Strains of SPCSV have also been identified in Argentina and Brazil and in Asia from their nucleic acid sequences (Winter et al., 1997). GEOGRAPHICAL ORIGIN OF SPCSV The SPCSV infects a wide range of wild Ipomoea spp. and a few non-Ipomoea spp. (Cohen et al., 1992). Sweetpotato originates from the Americas, was first domesticated c. 8,000 to 6,000 B. C. and was cultivated widely by pre-Columbian Amerindian civilisations by 2,500 B. C. (Austin, 1987). It may also have been taken to New Zealand, Papua New Guinea and Irian Jaya (formerly West New Guinea) (Yen, 1982). Post-Columbus, sweetpotato was also taken eastward, first to Spain and Portugal to become a luxury food in Europe (Zuckerman, 1999), and then to both West and East Africa and Asia in the 1500s (Yen, 1982; Austin, 1987). The genus Ipomoea and closely related members of the Convolvulaceae, some of which also seem likely to be infectible by SPCSV, are distributed worldwide, and these and Amaranthus spp. naturally host SPCSV (Milgram et al., 1996). Therefore, pre-Columbus, there seem to be three broad options for the distribution of SPCSV and its component strains, (i) the Americas, probably in sweetpotato or/and wild Ipomoea spp. and (ii) the ‘Old World’, probably in wild Ipomoea spp. and lastly worldwide, probably in sweetpotato or/and wild Ipomoea spp. There is no evidence that SPCSV is seed-borne but sweetpotato has tuberous roots that can survive many months, allowing them to be transported long distances. Tubers produced by infected plants carry SPCSV (Hahn et al., 1981) and readily produce adventitious shoots from which infected plants arise. So even casual disposal of tubers, for example, carried as food during a voyage, could result in new infected plants becoming established in distant locations and acting as foci of infection. If SPCSV has been transferred between the Old and the New World post-Columbus, infected tubers would seem to be its most likely means of transmission. Post-Columbus trade routes. In the 16 and 17th centuries, boats sailing to Europe from the Americas generally sailed there directly, carrying mainly primary products (sugar, cotton, metals, etc). For the return journey to the Americas, ships first sailed south to encounter easterly winds in equatorial regions, often landing in West Africa to exchange European trade goods for slaves to take to the Americas (Landes, 1998). To travel eastwards from Europe to the rest of the World prior to the opening of the Suez Canal, ships had to go south around Africa. Through these voyages, the SPCSV could have reached Europe from the Americas along with the crop itself and could have been carried to the rest of the world in vegetative planting material. Alternatively, if SPCSVWA originated in West Africa, the establishment of sweetpotato as a crop in Africa in the 16th Century (Yen, 1982; Austin, 1987) and its extensive cultivation in the Gold Coast (now Ghana) (Doku, 1984) would have allowed opportunity for infected sweetpotato tubers to be transported from there as stores on ships. Consequently, the presence of SPCSVWA in U. S. A. and elsewhere in the Americas could have resulted from such tubers from West Africa being discarded in the Americas. The presence of SPCSVEA in South America could similarly derive from Portugal’s early links with Mozambique and Brazil. Evidence of such long distance movement of sweetpotato is perhaps that sweetpotato cultivars found in Central Chile seem to have a non-Andean origin within recent historical time (Sagredo et al., 1998). Host plant reaction. Virtually all exotic cultivars and seedlings obtained by crossing amongst exotic clones of sweetpotato succumbed to SPVD when exposed at Namulonge Agricultural and Animal Production Research Institute, which is sited in a high SPVD incidence area in Uganda (Carey et al., 1997). This indicates that high levels of resistance to this disease are rare or absent in germplasm obtained from outside Africa. Furthermore, no single gene or genes for immunity to SPCSV have been reported in sweetpotato, unlike the situation with the Solanum potato for several of its sympatric viruses (Valkonen, 1994). Indeed, ‘New World’ wild species of sweetpotato such as I. setosa and I. nil are the preferred indicator plants for SPCSV because of their sensitivity, whereas several ‘Old World’ wild Ipomoea spp. are either very resistant to or tolerant to SPCSV (Karyeija et al., 1998b). These results suggest that neither sweetpotato nor its wild relatives in the New World had been exposed to much selection for resistance to SPCSV prior to arrival in the Old World but that Old World, wild Ipomoea species have long been exposed to SPCSV infections. Vector. The most efficient vector of SPCSV, B. tabaci, originated in the Old World, possibly Africa (Campbell et al., 1995), along with the other main whitefly-borne genus of viruses, the begomoviruses (Brown, 2000). Consistent with this, whiteflies were not reported on sweetpotato until 1982 in Peru and became common only in 1997 with the arrival of the B biotype (= B. argentifolii) of B. tabaci (Cisneros and Mujica, 1999), supporting an ‘Old World’ origin of SPCSV (Karyeija et al., 1998a). The SPCSV has recently been shown also to be transmitted, though less efficiently, by the banded-winged whitefly T. abutilonea (Sim et al., 2000). Phylogenetic relationships. Amongst eight other criniviruses including Lettuce infectious yellows virus (LIYV), the CP amino acid sequence of SPCSV was most similar to that of cucumber yellow stunting disorder virus (Livieratos et al., 1999), a virus apparently restricted to the Mediterranean Region and the Middle East (Wisler et al., 1998). However, the next most similar virus was LIYV, a virus with which SPCSVEA also shows a distant serological relationship (Hoyer et al., 1996) and which has been reported only from North America (Wisler et al., 1998). None of the above evidences precludes SPCSV originating in the New World but the balance of evidence, particularly the reactions to SPCSV of various Ipomoea spp. including sweetpotato, seems in favour of it having an Old World origin. Interestingly, the presence of SPCSVEA only in East and Southern Africa is mirrored by the distribution of its partner virus, SPFMV, a single distinctive cluster of SPFMV isolates having been found in East Africa (Kreuze et al., 2000). Consequently, this suggests that for both virus species either there was a restricted introduction into East Africa, or they evolved in East Africa from populations infecting the native flora. These hypotheses may be compared more precisely for both SPCSV and SPFMV when isolates of both species from a wider geographical range have been sequenced and their likely evolutionary relationships assessed. PARTICLE STRUCTURE, GENOMIC CONSTITUTION AND GENOMIC VARIABILITY OF SPCSV The SPCSV, a member of the Closteroviridae (reviewed by Karasev, 2000), belongs to the whitefly-borne genus, Crinivirus. While the LIYV (reviewed by Falk and Klaassen, 1995; Klaassen et al., 1995) is the type member of the genus. Closteroviruses are generally restricted to the phloem where intracellular inclusions including characteristic vesicles are commonly found (Duffus, 1995). Closteroviruses have long flexuous particles on which cross-banding may be visible under the electron microscope. The heat shock protein (70) (HSP70) homologue gene occurs uniquely in the Closteroviridae. Unlike members of the aphid-transmitted genus Closterovirus, which have a single ssRNA genome, the genome of criniviruses comprises two ssRNAs encapsidated in separate particles. In criniviruses, both the HSP70 homologue gene and the capsid protein (CP) gene occur in the smaller RNA 2 (Dolja et al., 1994). One function of the HSP70 homologue gene in closteroviruses may be to facilitate cell-to-cell movement in host plants (Agranovsky et al., 1998). The particles of SPCSV are 800 -920 nm long (modal length 850 nm) with a diameter of c. 12 nm and they have the open helical structure typical of closteroviruses. Membranous vesicles occur in phloem cells of infected I. setosa (Winter et al., 1992b). The bipartite nature of the genome of SPCSV (Anon., 1993) was confirmed by sequence analysis (Vetten et al., 1996). Hoyer et al. (1996) cloned and sequenced the CP gene of an isolate of SPCSV from Kenya. Its sequence resembled those of other members of the Closteroviridae including the aphid-borne Beet yellows virus (BYV) and Citrus tristeza virus (CTV), but especially that of another crinivirus LIYV. Polyclonal antibodies (Pabs) raised against CP obtained by expressing the SPCSV CP gene in Escherichia coli showed by immuno-sorbent electron microscopy (ISEM) plus decoration that sweetpotato closterovirus isolates from Israel, U. S. A., Nigeria and Kenya were all SPCSV (Hoyer et al., 1996). Significantly, only the isolate from Kenya was recognised in ISEM by an antiserum to LIYV. Monoclonal antibodies (Mabs) raised against the expressed CP of the Kenyan isolate of SPCSV and against partially purified preparations of an Israeli isolate of SPCSV enabled DAS- and TAS- ELISA to confirm that the Kenyan isolate was distinct serologically from the isolates from Israel, U. S. A. and Nigeria (Hoyer et al., 1996). Individual Mabs have further distinguished serotypes within SPCSVEA isolates from East Africa (Alicai et al., 1999a). Reverse transcription polymerase chain reactions (RT-PCR) utilising a primer pair located within the HSP 70 gene homologue of RNA 2, followed by single-strand conformation polymorphism analysis on the amplified PCR product, has also been used to differentiate isolates of SPCSV (Winter et al., 1997). Multiple sequence alignment analysis showed that SPCSV isolates from West African countries, USA and Israel form a cluster (presumably SPCSVWA) well separated from SPCSV isolates from Taiwan. Another cluster was formed by SPCSV isolates from Brazil and Argentina (Winter et al., 1997). These latter isolates were less than 85% identical to virus isolates of the other clusters. DETECTION OF SPCSV Adequate methods for detecting SPCSV, especially ones robust enough to work in the developing countries where SPCSV mostly occurs, have been developed only relatively recently (Gibson et al., 1998b). This delayed international appreciation of the importance of this virus. Even in the late 1980’s, grafting to a plant of a sweetpotato clone (Tib 8, s.c.9) highly susceptible to SPVD and infected with SPFMV was the recommended method for detecting SPCSV. Scions infected with SPCSV induced SPVD in the Tib 8 (Thottappilly and Rossel, 1988). Some important constraints to the development of serological methods of detection have been:
These constraints have been overcome in the 1990s in several ways. The first Pab to SPCSV was produced using SPCSVWA from Israel purified from infected I. setosa (Cohen et al., 1992). This antiserum could trap and decorate SPCSV particles specifically in ISEM. In order to produce larger quantities of antiserum and without anti-plant background reactivity, a polyclonal antiserum was raised subsequently in rabbits against antigen prepared using a bacterially-expressed coat protein gene of SPCSVEA (Hoyer et al., 1996; Vetten et al., 1996). The purified preparations of an Israeli isolate of SPCSVWA (Cohen et al., 1992) and the bacterially-expressed coat protein of SPCSVEA were also used to raise monoclonal antibodies (Mabs) (Vetten et al., 1996). These Pabs and Mabs have allowed the development of TAS-ELISA capable of identifying readily in sweetpotato infected with either SPCSVWA and SPCSVEA (Gibson et al., 1998b). These tests have been used in several developing countries including Africa. Because of the difficulties in purifying SPCSV, methods for detecting SPCSV nucleic acids were developed more-or-less contemporaneously with serological methods. The SPCSV RNA was reverse transcribed and cloned to produce SPCSV cDNA (Winter et al., 1992a). This reacted specifically in dot blot hybridizations with sap extracts from SPVD-affected sweetpotato, providing the first molecular method for detecting SPCSV. The cDNA was also used to make ssRNA probes (riboprobes) labelled with the steroid hapten, digoxigenin-UTP, during transcription. These labelled probes were subsequently detected using an alkaline phosphatase-conjugated anti-digoxigenin antibody and a chemiluminescent substrate (Winter et al., 1992a). This test took two days and was considered to be both more reliable and less time-consuming than bioassays for SPCSV using sweetpotato pre-infected with SPFMV: The International Board for Plant Genetic Resources (IBPGR), Rome, provided freely the cDNA clones (Anon., 1993). Winter et al. (1997) also used a RT-PCR-based method of detecting SPCSV based on a primer pair located within the heat shock protein (HSP 70) gene homologue. Single-strand conformation polymorphism analysis done on the amplified PCR product was used to distinguish SPCSV isolates from different continents. The RT-PCR was superior to the RNA dot-blot hybridisation, faster than grafting to the indicator plant, I. setosa, and could work with in vitro cultured sweetpotato, making it appropriate for quarantine purposes. However, Sim and Valverde (1999) considered that RT-PCR detection of SPCSV from sweetpotato using primers to the HSP70 homologue gene gave inconsistent results. SYMPTOMS OF SPCSV IN SWEETPOTATO AND IN INDICATOR SPECIES Host range. Criniviruses generally have limited host ranges (Duffus, 1995). Cohen et al. (1992) showed that, in addition to sweetpotato, SPCSVWA (= SPSVV) was transmitted by B. tabaci from I. setosa to: I. acuminata, I. hederacea, I. hederifolia, I. nil cv Scarlet O’Hara, I. purpurea, I. setosa, I. trichocarpa, I. trifida, I. wrightii, Nicotiana clevelandii, N. benthamiana and Amaranthus palmeri. On the contrary, SPCSVWA was not transmitted by B. tabaci from I. setosa to: Brassica pekinensis, Chenopodium murale, C. quinoa, Cucumis sativus, Datura stramonium, Daucus carota, Helianthus annuus, Lactuca sativa, Lycopersicon lycopersicon, Petunia hybrida, Physalis floridana, Raphanus sativus and Zinnia elegans. In addition, SPCSVWA (= SPVCV) was also transmitted by grafting to I. mexicana and I. Bona nox but not to N. tabacum, N. glutinosa, peanut or cotton (Loebenstein and Harpaz, 1960). Some perennial wild Ipomoea spp. such as I. cairica, I. involucra and I. wightii were not infected by graft inoculation with SPCSVEA; I. hildebrandtii was infected but was tolerant (Karyeija et al., 1998a). The SPCSV (= SPSVV) has also been identified in lisianthus (Eustoma grandiflorum) (Cohen et al., 2001). Symptoms. Criniviruses are generally restricted to the phloem and may cause microscopic changes including necrosis in this tissue. Visible symptoms generally comprise yellowing and/or vein yellowing in leaves of host plants and overall plant stunting (Duffus, 1995). Consistent with this, SPCSVEA from Uganda causes purpling or yellowing, depending on the cultivar, of the lower and middle leaves and some overall stunting of sweetpotato plants (Gibson et al., 1998a) (Plate 1). By contrast, SPCSVWA from Nigeria, Israel and U.S.A. is reported to be symptomless in a range of sweetpotato cultivars (Schaefers and Terry 1976; Winter et al., 1992b; Sim and Valverde, 1999). Infection by SPCSVEA caused seedlings of the indicator plant, I. setosa, to become severely stunted and for the leaves to become small, brittle, yellow and rolled downward (Hoyer et al., 1996; Gibson et al., 1998a). Impomea setosa infected with SPCSVWA were also stunted with small, brittle leaves (Cohen et al., 1992), but had inwardly-rolled leaves (Winter et al., 1992b). Atypically for a crinivirus, SPCSV from Argentina identified serologically as SPCSVWA induced a mild mosaic in both I. setosa and N. benthamiana and very mild mosaic in old and new leaves of sweetpotato. Infection of sweetpotato with both SPCSV and SPFMV is required to cause SPVD (Schaeffers and Terry, 1976). Stunting and chlorosis of the leaves, sometimes combined with a purpling seen in SPVD-affected plants are typical symptoms induced by a crinivirus; the chlorotic mottle or vein clearing and distortion of leaves also seen in affected plants are typical of a potyvirus. However, symptoms of SPVD vary considerably. Leaves of varieties which are deeply dissected often become extremely so, such that the leaves appear strap-like (Plate 2). Cultivars can develop such mild SPVD symptoms that farmers cannot readily distinguish between affected and unaffected plants (Thottappilly and Rossel, 1988). Isolates of SPCSVEA differing in the severity of associated SPVD have also been identified and were distinguishable by their reaction to specific Mabs (Alicai et al., 1999a). Co-infection of I. setosa with SPCSV and SPFMV usually causes very severe symptoms. Ugandan isolates of SPCSVEA and SPFMV caused very severe stunting, chlorosis and necrosis of young leaves in I. setosa (Gibson et al., 1998a). The SPCSVWA and SPFMV from Nigeria initially produced stunting and chlorotic leaf symptoms and severe epinasty in the subsequent chronic phase of infection (Winter et al., 1992) and the older leaves became necrotic (Schaefers and Terry, 1976). Unfortunately, these observations of slightly different symptoms are confounded with differences in experimental conditions, the co-infecting isolate of SPFMV and/or plant genotype, and no side-by-side comparisons of the symptoms of SPCSVEA and SPCSVWA isolates have been reported. YIELD EFFECTS OF SPCSV AND SPVD Sweetpotato virus disease is the most important disease of sweetpotatoes globally (Carey et al., 1999), having a major effect on tuberous root yields (Tables 1 and 2). All reported trials involve plots planted wholly either with cuttings taken from unaffected or diseased plants (Table 1) or with inoculated and virus-free plants (Table 2). However, in farmers’ fields, most cuttings are obtained from unaffected parents and infection occurs during growth (Gibson et al., 2000). Furthermore, only a proportion of plants is usually affected, allowing opportunity for unaffected neighbours to compensate by increased growth. Consequently, despite the dramatic yield effects recorded in Tables 1 and 2, the actual effects of SPCSV or SPCSV + SPFMV (= SPVD) may be relatively minor in farmers’ fields until incidence exceeds 50% (Aldrich, 1963). However, SPCSV also has a major indirect effect on yield of sweetpotato in Africa. This effect is because most resistant cultivars currently used by African farmers have a relatively poor yield (see section on Resistance ahead) and the inability of farmers to grow high yielding susceptible cultivars in areas where SPVD is prevalent seems likely to be a considerable constraint on sweetpotato production (Aritua et al., 1998b). Furthermore, the failure of any high-yielding exotic sweetpotato varieties to be released in Uganda (the main producer in Africa) is largely the result of their susceptibility to SPVD and the prevalence of this disease. SYNERGISTIC EFFECTS OF SPCSV ON OTHER CO-INFECTING VIRUSES The first reported evidence of the synergistic effects of SPCSV on other viruses was that SPFMV could be acquired readily only from sweetpotato plants infected with both SPFMV and SPCSVWA (Schaefers and Terry, 1976). Similar results were obtained in Uganda using SPFMV and SPCSVEA (Aritua et al., 1998a). The titre of both coat protein and viral RNA of the SPFMV was increased (hundredfolds) by co-infection (Gibson et al., 1998a; Karyeija et al., 2000a) (Table 3). However, the titre of SPCSV remained similar in both plants infected with SPCSV alone or also with SPFMV. The increased titre of SPFMV in sweetpotato infected additionally with SPCSV is consistent with the mosaic or vein clearing symptoms associated with SPVD being typical of those of a potyvirus rather than of a crinivirus (Aritua et al., 1998a). Thus, SPVD appears to be the result of infection with SPCSV leading to the breakdown of resistance to SPFMV. Infection with SPCSV can break even the extreme resistance of the sweetpotato clone 420020 to SPFMV infection (Karyeija et al., 2000b). How it does so remains the subject of research. The SPCSVEA remained restricted to the phloem of sweetpotato whereas the titre of SPFMV in dually-infected plants seems to be raised throughout the plant (Gibson et al., 1998a; Karyeija et al., 2000a). The rate of long distance movement of SPFMV within sweetpotato plants was also unaffected by the presence or absence of SPCSV. The SPCSV may enhance the multiplication of SPFMV in tissues other than where it itself occurs by interfering with the systemic signalling required to maintain resistance to SPFMV (Karyeija et al., 2000a). This SPFMV-resistance-breaking attribute of SPCSV is clearly key to the development of SPVD and breeding SPVD-resistant sweetpotato is dependent on developing cultivars resistant to SPCSV. SPCSV, together with SPFMV and another potyvirus Sweetpotato mild speckling virus (SPMSV) causes sweetpotato chlorotic dwarf disease, a particularly severe disease of sweetpotato in Argentina. The SPCSV also acts synergistically with SPMSV alone to produce more severe symptoms than either virus alone. The SPMMV has also been found associated with SPCSV and SPFMV (Gibson et al., 1998a); perhaps this potyvirus also benefits from co-infection with SPCSV. Interestingly, the ability of SPCSV to synergise infection of sweetpotato also extends to Cucumber mosaic virus (CMV: Cucumoviridae). Thus, although virus-free sweetpotato cv Georgia Jet is immune to CMV, it is susceptible when pre-infected with SPCSV (Cohen and Loebenstein, 1991). TRANSMISSION OF SPCSV Criniviruses are usually transmitted by Bemisia spp. or Trialeurodes spp., any one crinivirus usually being transmitted by whitefly species of only one genus (Rubio et al., 1999). T. vaporariorum, T. abutilonea and B. tabaci have each been shown to transmit at least one crinivirus. Criniviruses are transmitted by whiteflies in the semi-persistent manner, persisting in their vector for no more than a few days. Acquisition depends on feeding rather than just probing, and generally reaches a maximum within a day or so (Duffus, 1995; Wisler et al., 1998). Criniviruses are not normally sap-transmissible (Wisler et al., 1998). Hansford (1944) first suggested an association between whiteflies Bemisia spp. and the spread of SPVD after observing many whiteflies in plots severed by SPVD. The SPCSV was shown experimentally to be transmitted by whiteflies in the late 1950s in Africa, perhaps under the synonym of virus B (Sheffield, 1957), in U.S.A. under the synonym(s) of sweetpotato mosaic and/or virus yellows (Girardeau, 1958; Hildebrand, 1960), and/or in Israel under the synonym of SPVCV (Loebenstein and Harpaz, 1960). However, Schaefers and Terry (1976) in Nigeria first reported the transmission by B. tabaci of what was definitely SPCSV. SPCSV is, as expected for a crinivirus, transmitted by whiteflies in the semi-persistent manner. B. tabaci can acquire or transmit SPCSV with access periods of 1 hr or less and can remain viruliferous for 24 - 48 hr (Larsen et al., 1991). Batches of 50 B. tabaci infected 16, 56 and 84% of I. setosa seedlings following, respectively, 1, 6 and 24 h inoculation access periods (IAP). Batches of 50 B. tabaci given 1, 6 and 24 h acquisition access periods (AAP) to an SPCSV-infected I. setosa infected 21, 59 and 88% of I. setosa seedlings, respectively (Cohen et al., 1992). SPCSV is transmitted moderately efficiently by B. tabaci. Thus, batches of 10, 25 and 50 B. tabaci given 24 hr AAP on an SPCSV-infected I. setosa infected 13, 31 and 94% of I. setosa seedlings during 24 h IAP. Bemisia tabaci is distributed worldwide and a recent pandemic of the B biotype (= B. argentifolii) in the Americas has been associated with an increase in whiteflies on sweetpotato, at least in Peru (Cisneros and Mujica, 1999), as has also occurred on many other crops worldwide. The A and B biotype can differ in their abilities to transmit specific criniviruses (Duffus, 1995). However, tests of whether the B biotype can transmit SPCSV have not been reported. In Africa, SPCSV can be transmitted by both the cassava-specific (Burban et al., 1992) biotype of B. tabaci (Schaefers and Terry, 1976) and B. tabaci naturally colonising sweetpotato (Cohen et al., 1992; Gibson et al., 1998a). B. afer has also been reported in Africa (Munthali, 1992) on sweetpotato (Legg et al., 1994); no tests of its ability to transmit SPCSV have been reported. Unusually for a crinivirus, SPCSV can also be transmitted by a whitefly in a second genus, the banded winged whitefly T. abutilonea, although less efficiently than by B. tabaci (Sim et al., 2000). Single T. abutilonea given 2 day AAP to infect sweetpotato and 2 day IAP to I. nil seedlings infected 4 out of 123 (3%) of the I. nil. However, in similar experiments, 18 out of 118 (15%) of I. nil inoculated with B. tabaci became infected with SPCSV. Both whiteflies occur in sweetpotato fields in U. S. A, although most criniviruses are only transmitted by either Bemisia spp. or Trialeurodes spp., Tomato chlorosis virus is also transmitted by both (Wisler et al., 1998). The SPCSV is graft-transmissible (Rossel and Thottappilly, 1988) but is not sap-transmissible (Larsen et al., 1991; Cohen et al., 1992). EPIDEMIOLOGY OF SPCSV Hansford (1944) linked the spread of SPVD to the prevalence of whiteflies of Bemisia spp. However, the first numerical association between whitefly populations on sweetpotato throughout the year and SPVD incidence was made 54 years later (Aritua et al., 1998b). In East Africa, whiteflies are especially abundant on sweetpotato during the hot dry period from November to May and especially on crops 2 – 4 months old (Sheffield, 1957; Wambugu, 1991; Aritua et al., 1998b; Alicai et al., 1999a). The SPVD requires infection by both the whitefly-borne SPCSV and the aphid-borne SPFMV, infection with the SPCSV enabling SPFMV to multiply rapidly (Karyeija et al., 2000b). In Nigeria, most plants of sweetpotato are infected with SPFMV, so super-infection with SPCSV automatically triggers the development of SPVD (Thottappilly and Rossel, 1988). In Uganda, local varieties of sweetpotato seem to be somewhat resistant to SPFMV when infected by this virus alone (Aritua et al., 1998a) and the majority of symptomless plants in farmers’ fields tested free of SPFMV (Gibson et al., 1997). Although no aphids were found on plants during monthly monitoring in farmers’ fields in Uganda (Aritua et al., 1998b), transmission of SPFMV is assumed to be by itinerant alate aphids (Wambugu, 1991). That the spread of SPVD has largely been associated with the abundance of whiteflies (Aritua et al., 1998a, 1999) suggests that inoculation with SPCSV rather than SPFMV is the limiting factor, suggesting that SPFMV is commonly present either latent in sweetpotato plants or in viruliferous aphids. Sheffield (1953) encountered differing incidences of SPVD on sweetpotato crops in Kenya, Uganda, Tanzania, Rwanda and Burundi and suggested that the disease was common at mid-altitudes. However, it was less common in districts on the eastern shore than on the western shore of Lake Victoria (Aritua et al., 1998b; Aritua et al., 2000; Gibson et al., 2000) (Fig. 1) despite these locations all being mid-altitude (c. 1,100 m above sea level). Prevailing winds ensure that rainfall is more evenly spread throughout the year on the western than on the eastern shore of Lake Victoria. The SPVD is also common at low altitude, along the Indian Ocean coast, particularly the relatively wet coast of Tanzania (Fig. 1). Despite whitefly populations peaking during the hot dry season (Aritua et al., 1998b) and declining during periods of rainfall (Alicai et al., 1999b), rainfall appears to have an overall beneficial effect on B. tabaci by promoting crop growth. Similar results have been found with B. tabaci on cassava (Legg, 1994). Rainfall throughout the year also encourages the growth of tall shrubs and trees, reducing ground level wind turbulence and providing a sheltered environment for whitefly to multiply on sweetpotato crops (Jeremiah et al., 2001). By contrast, north-eastern Uganda, where whiteflies and SPVD are relatively rare (Aritua et al., 1998b), has a long dry season in which vegetation is frequently burnt. The vegetation is consequently largely grassland, allowing strong winds to occur at crop level. In Israel, SPCSV spreads quickly to isolated plots of virus-free sweetpotato, and Amaranthus spp. and several species of Convolvulaceae may be natural reservoirs (Milgram et al., 1996); natural infections have also been recorded there in I. purpurea and I. mexicana (Loebenstein and Harpaz, 1960) and in crops of lisianthus (Cohen et al., 2001). Elsewhere, sweetpotato crops seem to be the main source of SPCSV. In Nigeria, plants isolated from diseased crops were rarely affected by SPVD (Thottappilly and Rossel, 1988). In Mpigi District in Uganda, the spread of SPVD into plots of two moderately resistant varieties of sweetpotato in farmers’ fields, correlated positively with the incidence of SPVD in nearby sweetpotato crops (Aritua et al., 1999). Surveys of the incidences of SPVD in Kagera district of Tanzania have shown a similar positive correlation between the incidence of SPVD and the proximity of the nearest sweetpotato crop. In Africa, sweetpotato is typically grown near homes, in small fields grouped close together and with new and mature crops overlapping or even the same field being used repeatedly for sweetpotato (Karyeija et al., 1998a), and this facilitating further of the disease spread. The vector (B. tabaci) occurs in at least two biotypes in Africa, one colonising cassava and the other colonising a range of other crops including cotton, various Cucurbitaceous and Solanaceous crops, and sweetpotato (Burban et al., 1992; Legg, 1996). The two biotypes differ in isoenzyme pattern (Legg et al., 1994), B. tabaci obtained from sweetpotato or cotton crops fails to develop if confined to cassava, and B. tabaci obtained from cassava barely survives on sweetpotato or cotton (Legg, 1996). Cassava is an important crop throughout much of the areas in Africa where sweetpotato is widely grown and is often grown with sweetpotato in the same smallholdings or the same fields. Furthermore, the cassava biotype of B. tabaci can transmit SPCSV (Schaefers and Terry, 1976). However, the unwillingness of the cassava biotype to colonise sweetpotato (Legg, 1996) seems to prevent B. tabaci on cassava being involved to any significant degree in the spread of SPCSV. This failure to colonise is currently important because a pandemic in East and Central Africa of the B. tabaci-transmitted cassava mosaic geminiviruses is associated with massively increased populations of B. tabaci on cassava (Legg, 2000). This is presumably associated with the lack of any reports of an associated increased spread of SPVD in sweetpotato. Crops of sweetpotato in eastern Uganda, where large numbers of whiteflies have been recorded on cassava (Legg and Ogwal, 1998), have low incidences of SPVD (Aritua et al., 1998b) despite intercropped cassava being devastated by cassava mosaic disease (Plate 3). RESISTANCE IN SWEETPOTATO TO SPVD The need for SPVD-resistant varieties of sweetpotato was evident in East Africa >50 years ago (Hansford, 1944), was unsatisfied two decades later (Aldrich, 1963) and still largely remains so (Gibson et al., 2000). Breeding for SPVD-resistance was first done at the International Institute of Tropical Agriculture (IITA) in Ibadan, Nigeria (Hahn et al., 1989). There, SPVD spread only slowly, susceptible plants taking 1-3 years exposure in the field to become diseased; raising vectors artificially and releasing them in the field did not give much increase in spread. Consequently, tuberous-root, core-grafting with tuber plugs taken from SPVD-affected tubers was adopted for screening sweetpotato for resistance to SPVD. The leaves of sweetpotato sprouts that arise from such grafted storage roots produce characteristic SPVD symptoms usually within 11 to 13 days. Severity of symptoms significantly (P<0.01) correlated with results of natural infection in fields. Typically, about 85-90% of sprouting core-grafted tubers of susceptible varieties were diseased compared with only 10% for a relatively resistant clone (TIS 2498). The core-graft method can, thus, be applied as a mass selection to screen sweetpotato breeding material for resistance to SPVD at an early stage in breeding. Heritability of resistance to SPVD was high allowing resistant seedlings to be identified accurately (Hahn et al., 1981; Hahn et al., 1989). Several resulting clones have been adopted in West Africa (Whyte, 1992). Varieties bred by the International Institute for Tropical Agriculture (IITA) in West Africa against SPVD caused by SPCSVWA and Nigerian isolates of SPFMV succumbed to SPVD when exposed at Namulonge Agricultural and Animal Production Research Institute (NAARI), Uganda (Mwanga et al., 1991) and elsewhere in East Africa. Presumably, this is because they were then exposed to SPCSVEA (Gibson et al., 1998b) and/or East African isolates of SPFMV (Kreuze et al., 2000). Furthermore, whilst early demand had been for SPVD-resistant varieties (Hansford, 1944, Aldrich, 1963), it became evident that SPVD-resistant landraces were available to farmers. These were being grown, especially in localities where B. tabaci was abundant (Aritua et al., 1998b). Although many farmers knew of resistant varieties, they also considered that most had undesirable characters such as poor and/or late yielders, poor taste and poor in-ground storability and contested that superior resistant varieties were one of their greatest needs (Gibson et al., 2000). East African farmers in areas where SPVD was prevalent were, consequently, making compromises in their choice of cultivars between good yields and adequate virus resistance (Aritua et al., 1998b) (Table 4). This shifted the emphasis to the identification of clones with good yields and other beneficial attributes, and which were also resistant to SPVD. The Potato Programme at NAARI first screened local Ugandan germplasm to identify high-yielding resistant landraces (Mwanga et al., 1991). Five of these landraces have now been released for national dissemination (Mwanga et al., 2001). An associated conventional breeding programme, based on crosses amongst superior local and introduced germplasm (Carey et al., 1997), has led to the recent release of a further six SPVD-resistant, high-yielding varieties in Uganda in 2000 (Hakiza et al., 2000). A major difficulty in the breeding work has been that very large progeny populations (as many as 100,000 seedlings) have been required to identify just a few SPVD-resistant, agronomically superior genotypes (Carey et al., 1999). This seems likely to be associated with resistance being polygenic which, though disadvantageous in breeding terms, may benefit the durability of resistance bearing in mind the diversity of SPCSV even within East Africa (Alicai et al., 1999b). The diversity of SPCSV in Africa may also explain why no immune clone of sweetpotato has been identified. In Uganda, SPVD has been observed to spread rapidly and unassisted spread of SPCSV and SPFMV by natural populations of whiteflies and aphids, respectively, has sufficed to screen for resistance. Unlike screening by core-grafting, this provides opportunity to select for vector resistance. Despite this, whiteflies were similarly abundant on clones of sweetpotato differing considerably in their resistance to SPVD (Aritua et al., 1998a; Alicai et al., 1999a), suggesting that there was no selection for vector resistance. However, even though SPVD-resistant varieties may have no additional controlling effect on whiteflies, the widespread cultivation of the resistant sweetpotato variety New Kawogo appears to be providing protection to adjacent crops of susceptible cultivars by limiting the numbers of viruliferous whiteflies that reach them (Aritua et al., 1999). Aphids, by contrast, generally seem to be absent from sweetpotato crops in East Africa; SPFMV is probably being transmitted by non-colonising itinerant aphids (Wambugu, 1991), the non-persistent manner of transmission of SPFMV circumventing vector resistance. Although the impact of varieties of sweetpotato bred on-station is likely to increase, most sweetpotato production in Africa is based on local varieties (landraces), of which there may be several hundred in any one region (Bashaasha et al., 1995; Kapinga et al., 1995). Little is known of the origin of these landraces. They may differ considerably in their resistance to SPVD, associated with the local prevalence of whiteflies on the crop (Aritua et al., 1998b), implying that SPVD has led to susceptible landraces being eliminated (Aldrich, 1963) or, perhaps more commonly, for susceptible seedlings to fail to become established. Despite the wide range of landraces present, farmers generally seem to know and care little about the survival of seedlings, new landraces apparently establishing with little or no assistance from research scientists (Gibson et al., 2000). Indeed, the main underlying cause for the continuing prevalence of SPVD in East Africa may be that the traditional sweetpotato farming system lacks an adequate system for breeding the high-yielding but resistant landraces required (Gibson et al., 2000). This has been exacerbated by very large progeny populations required to identify just a handful of resistant, agronomically superior genotypes (Carey et al., 1999). The success, albeit limited, of landraces suggests that a little assistance, such as educating farmers to appreciate the potential value of seedlings and perhaps supplying them with seedlings, could help facilitate identification of superior landraces rapidly. Sweetpotato clones resistant to SPVD seem mostly to have resistance of a type that restricts the number of plants that develop SPVD (Aritua et al., 1998a; Alicai et al., 1999). However, clones may also differ in response to infection (Ngeve and Boukamp, 1991) and all plants of tolerant cultivars such as the West African cultivar Dukukpuku may be so mildly affected by SPVD that farmers cannot readily discriminate against taking cuttings from affected plants (Thottappilly and Rossel, 1988). The yield penalty of a mild infection of all plants in a cultivar may be greater than the yield penalty of a few severely diseased plants, especially since the sprawling indeterminate growth of most cultivars allows unaffected neighbouring plants to compensate effectively (Aldrich, 1963). Perhaps because of this, clones in which all plants are mildly affected are rare, at least in East Africa (Gibson et al., 1997). The SPVD is, consequently, obvious in most clones, allowing farmers to avoid using them in future plantings (Bashaasha et al., 1995; Kapinga et al., 1995; Gibson et al., 2000). If so, this suggests that, in the absence of immunity, the ideal resistant cultivar is one which is seldom infected but develops obvious symptoms when infected. A few clones of sweetpotato may also be so resistant to SPVD that they recover, producing unaffected foliage (Hahn et al., 1981; Terry, 1982). It is unclear whether cuttings taken from such foliage remain symptomless and are free of SPCSV and SPFMV. In cassava, recovery and reversion are key elements in enabling the sustained availability of planting material free of cassava mosaic disease in African smallholder farmers (Fargette et al., 1994). Since SPVD is caused by a dual infection with SPCSV and SPFMV, resistance to the disease could be the result of resistance to SPFMV, SPCSV or a mixed infection of these diseases. However, symptoms of SPCSV alone are the first sign of the eventual development of SPVD both in artificial and field infections (Gibson et al., 1998b) and without pre-infection with SPCSV, SPFMV has difficulty in infecting at least some East African sweetpotato genotypes (Gibson et al., 1997; Aritua et al., 1998a). This suggests that resistance to SPCSV is the key feature of the current SPVD-resistant cultivars. So far, it has been largely the African sweetpotato gene pool that has been searched for sources of resistance. The apparently increasing incidence of SPCSV in South America (Carey et al., 1999; Cisneros and Mujica, 1999; Di Feo, 2000) may identify better sources of resistance from within the much wider American sweetpotato gene pool. In Africa, wild species such as I. cairica, I. involucra and I. wightii are not infected by SPCSV and I. hidebrandtii is tolerant (Karyeija et al., 1998b) but transfer of resistance from such species into I. batatas is hindered by genetic incompatibility. The coat protein (CP) of SPFMV has been expressed in sweetpotato and provided a heritable form of resistance (Okada et al., 2001) to diverse isolates of SPFMV, suppressing virus titre even following graft inoculation (Nishiguchi et al., 2001). Utilising the SPCSV CP or another viral gene may be an alternative sources of resistance to SPCSV too. Such engineering of resistance would have the advantage that it could be done on cultivars already established as high yielding. CULTURAL CONTROL PRACTICES Sweetpotato is grown in a continuous cycle of vegetative production in most African countries, crops from one planting season overlapping with another, and cuttings obtained from mature crops being used to plant succeeding crops with sometimes two or even three main planting seasons each year. Piecemeal harvesting of individual tubers is common and extends the cropping season, even up to a year after crop maturity (Bashaasha et al., 1995; Kapinga et al., 1995). Even where there is a prolonged dry season, planting material still has to be maintained vegetatively, for example, in shaded or in low-lying marshy areas, or as dormant tubers. This continuous cycle of vegetative production “provides a highly efficient mechanism for the perpetuation and dissemination of sweetpotato viruses” (Terry, 1982) including SPCSV. There seems no obvious way of breaking this cycle but some cultural interventions may facilitate control of SPVD. Examples of such cultural practices include selection of disease-free planting material, destruction (roguing) of diseased plants, especially in young crops, isolating new crops from old diseased crops, and destroying crop residues. Sweetpotato may also be intercropped, often with fast-growing crops such as maize and beans (Kapinga et al., 1995); intercropping, especially with maize, may provide some control of SPVD through decreasing vector numbers due to unfavourable micro-climate (Ndunguru and Aloyce, 2000). Selection of planting material. Farmers generally select cuttings for new crops preferentially from plants unaffected by SPVD (Bashaasha et al., 1995; Kapinga et al., 1995; Aritua et al., 1998b) and regard this as their main means of controlling SPVD (Gibson et al., 2000) (Table 5). It maintains an equilibrium level of unaffected plants, selection counterbalancing spread by whiteflies (Thottappilly and Rossel, 1988). Indeed, cuttings selected from symptomless sweetpotato plants of traditional cultivars in East Africa have been found to be mostly virus-free, yielding similarly to cuttings taken from plants known to be virus-free (Carey et al., 1999). Selection of cuttings from unaffected plants depends on infected plants having obvious symptoms and tolerant cultivars that express inconspicuous symptoms may become completely infected (Thottappilly and Rossel, 1988). Specifically grown virus-free planting material can provide large yield gains, especially in commercial, and relatively virus susceptible, varieties (Carey et al., 1999). Its use has been shown to be highly profitable in China (Fuglie et al., 1999; Gao et al., 2000). It is also being used by commercial farmers in southern Africa (Joubert et al., 1979) and Egypt (Carey et al., 1999). In South Africa, new stock is introduced every three years (Meynhardt and Joubert, 1982). Increased commercialisation of the crop may result in the increased use of such material elsewhere in Africa. Destruction. Although farmers usually select cuttings preferentially from symptomless parents, some SPVD-affected plants often occur even in young crops. Some farmers remove diseased cuttings in young crops (Gibson et al., 2000) (Table 5), a practice which has been long recommended (Aldrich, 1963). However, most farmers do not rogue, probably due to a misconception that removing the young plants will cause a proportional loss in yield. However, most cultivars of sweetpotato have a sprawling, indeterminate growth habit and unaffected neighbours can make up, often by an increase in the proportion of large tubers (Schultheis et al., 1999), most of the small yield which diseased plants might otherwise have produced. Thus, in Uganda, plants severely affected by SPVD produced only 33% of the yield of unaffected plants, whereas halving planting density changed yield by only c. 10%. It was, therefore, suggested that removing diseased plants may not seriously affect yield as long as no more than 50% of plants are removed (Aldrich, 1963). Farmers may also not rogue because they do not consider the fact that SPVD-affected plants represent a source of infection for other plants, commonly assuming that SPVD has an abiotic cause (drought, too much sun, poor soil, etc). Such an assumption is not unusual (Bentley, 1991) because weather often triggers disease outbreaks: small-scale potato farmers in South America and Uganda also consider that late blight due to Phytopthora infestans is caused by the bad weather (Ortiz et al., 1999) that favours the pathogen. Destroying old crops. The ease with which tuberous roots can develop shoots and foliar cuttings can root results in diseased sweetpotato often surviving in harvested fields. Plate 4 shows such an unharvested tuber sprouting with SPVD-affected foliage. Since sweetpotato can also yield in relatively nutrient depleted soils, it is often grown as the last crop before a fallow so the field is then abandoned (Bashaasha et al., 1995) with no subsequent weeding in preparation for a new crop. It may be difficult to persuade farmers to remove sweetpotato from abandoned crops but grazing by domestic animals may be an acceptable control. Isolating new crops. Sweetpotato is often planted throughout the year so as to provide a continuous supply of fresh tubers and the practice of piecemeal harvesting extends cropping. Consequently, new and mature crops overlap. Sweetpotato is commonly grown in Africa in small fields, planted near homes and new crops of sweetpotato are often planted close to or even directly adjacent to old crops (Karyeija et al., 1998a) (Plate 5). Since spread of SPCSV is closely related to the proximity of diseased sweetpotato crops (Aritua et al., 1999), even small increases in distance of new plantings from old crops should provide worthwhile benefits. Again, failure to realise that SPVD is a disease spread by whiteflies may prevent farmers appreciating the benefits of planting new crops as far away as possible from old ones. Furthermore, although it may be inconvenient, unlike roguing, isolating new crops has no inevitable yield losses unlike plant removals. Phytosanitation provides control options, which work in a ‘virtuous’ cycle, roguing and isolation reducing virus incidence in mature crops, so making it less likely that cuttings taken from them are latently infected and so fewer cuttings subsequently need to be rogued. Phytosanitation also integrates well with the use of resistant varieties (Terry, 1982) and besides, isolation of new crops and destruction of crop debris as a means of controlling SPCSV may also assist in the control of sweetpotato weevils (Cylas spp.), the main pest of the crop in much of Africa. However, the evidence for economic benefits may need to be more tangible if farmers are to be persuaded of its benefits. One of the main benefits may be that phytosanitation can enable high yielding or otherwise superior susceptible varieties to be grown (Aritua et al., 1998b; Gibson et al., 2000). Phytosanitation may also have the long-term benefit of maintaining in use the widest possible range of genetic material (Aldrich, 1963). However, if farmers are to adopt and practise phytosanitary methods successfully, they will need to be better informed of the true cause and means of spread of SPVD. CONCLUSIONS Despite considerable research on SPCSV, a lot of aspects about the virus remain unknown. The precise distribution of SPCSVEA and SPCSVWA, where they overlap and how they interact there are unknown. The distribution of these strains outside Africa is not clearly understood. The SPCSV has been reported in Taiwan, but not in mainland China, S. E. Asia or India although its presence seems likely. There are also indications that strains other than SPCSVWA and SPCSVEA may occur worldwide (Winter et al., 1997). It is clearly important both for Africa and elsewhere that SPCSV is not distributed more widely. The latter may be difficult in sub-Saharan Africa where there are mainly land borders between countries but the apparent ability of SPCSVEA to damage sweetpotato varieties bred in West Africa with resistance to SPCSVWA (Mwanga et al., 1991) highlights the need to restrict the spread of these different strains. It is not known whether the two strains can co-infect sweetpotato; if they can, dual infection may cause a more severe disease infestation than those caused by either strain infecting separately. Knowledge of the exact distribution of the two strains is needed to facilitate their containment. Improved knowledge of the distribution of SPCSV may also allow the identification of locations where more resistant cultivars may be found and knowledge of the worldwide distribution of different strains of SPCSV seems to be essential if resistance is to be exploited efficiently. Thus, East African sweetpotato varieties may provide a valuable source of resistance wherever SPCSVEA occurs. Remarkably little is known about the epidemiology of SPCSV; studies have been reported only from East Africa and Israel. This is an important omission because preliminary evidence suggests that phytosanitation can provide valuable control. That most spread is local (Aritua et al., 1999) is consistent with SPCSV being transmitted semi-persistently. Local inoculum may be reduced by planting disease-free cuttings (already used), roguing (used by a few farmers early in the season), removal of crop debris (both roots and foliage as both re-grow readily) from old fields as this is often diseased (not used), temporal or spatial separation of new crops from old diseased ones (not used), and increased use of resistant varieties (used but favoured only in localities where there were acceptable resistant varieties). However, despite the obvious merits of these approaches, none has yet been tested in field trials. That most farmers have incorrect concepts of the cause and source of SPVD is also likely to cause difficulties as they may imply to farmers that such strategies are inappropriate. Phytosanitation requires the active participation of farmers and consequently, both evaluation and teaching programmes will be essential components if such practices are to be utilised. Land use, land ownership and other socio-economic issues are also likely to be important issues in phytosanitation: men generally have the most power over land but, in Africa, sweetpotato is mainly a woman’s crop (Bashaasha et al., 1995; Kapinga et al., 1995). Finally, the synergism by which SPCSV enables a range of viruses to damage sweetpotato currently presents perhaps the most fascinating scientific aspect of this virus. Elucidating the mechanism (Karyeija et al., 2000b) may provide novel means of controlling SPVD and may also provide additional information of how viruses invade plants. ACKNOWLEDGEMENTS Preparation of this manuscript was supported by Project R7492 of the UK DFID Crop Protection Programme and Project ICA4-CT-2000-30007 of the EU INCO-DEV Programme. The views expressed are not necessarily those of these Programmes. The authors thank Prof. J.M. Thresh, Dr L. Kenyon and an anonymous referee for critically reviewing the manuscript. REFERENCES
©2002, African Crop Science Society The following images related to this document are available:Photo images[cs02028t2.jpg] [cs02028f1.jpg] [cs02028t4.jpg] [cs02028p2.jpg] [cs02028t5.jpg] [cs02028p4.jpg] [cs02028p5.jpg] [cs02028t1.jpg] [cs02028t3.jpg] [cs02028p3.jpg] [cs02028p1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
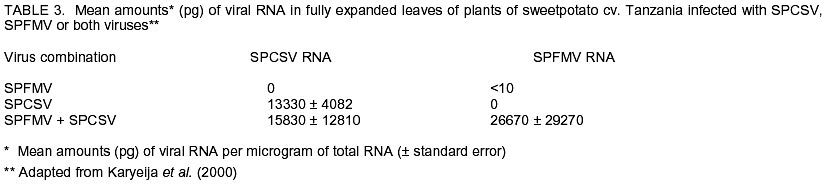{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}