 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
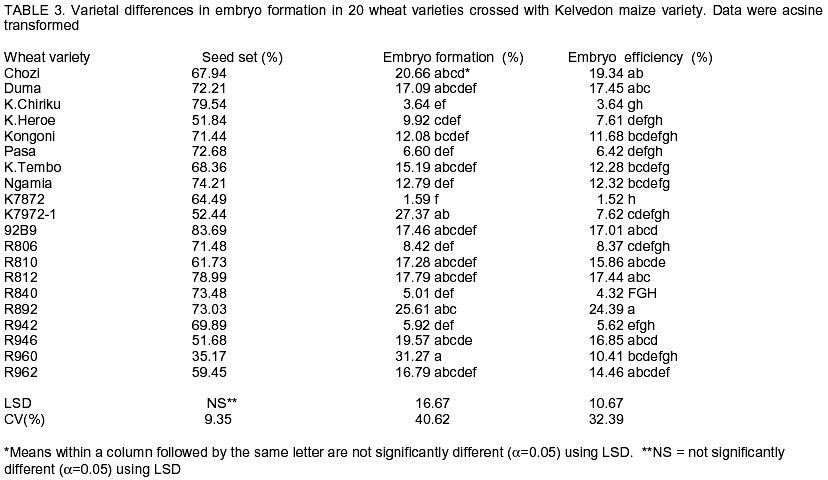
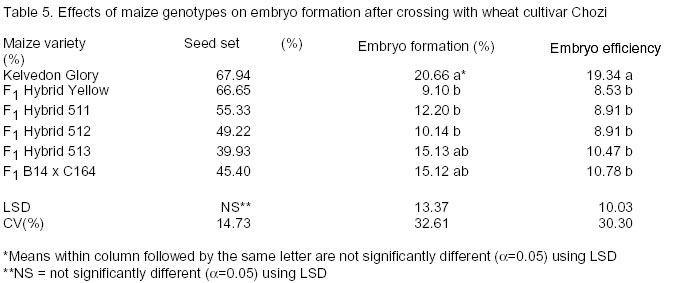
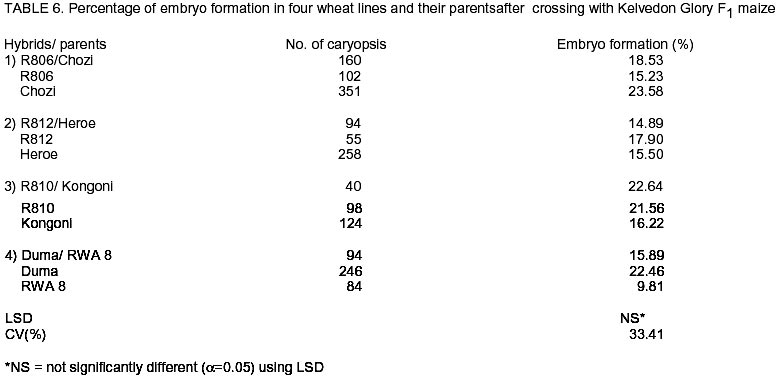
African Crop Science Journal, Vol. 10. No. 4, 2002, pp. 311-316 Development of a doubled haploid system for wheat through wheat x maize crosses in Kenya L. Karanja, M. Kinyua and J. Maling’a National Plant Breeding Research Center, Njoro, P.O. Njoro, Kenya (Received 12 October, 2001; accepted 15 July, 2002) Code Number: cs02029 Abstract Doubled haploid technique facilitates the development of homozygous plants within one generation, tremendously reducing breeding time. Although the system has been successfully established in Kenya, improvement of protocol efficiency remains a critical and potentially limiting factor. The objective of this study, therefore, was to investigate wheat (Triticum aestivum) and maize (Zea mays L.) genotype effect on efficiency of haploid production aimed at establishing an appropriate protocol for Kenyan conditions. Twenty wheat genotypes were crossed with six maize varieties. The haploid embryos were rescued and cultured for plant regeneration and subsequently treated with colchicine for chromosome doubling. Half- diallel crosses were made in a cage and greenhouse and the embryos were cultured in the laboratory under controlled environment. Significant genotypic differences in seed set (51.6-90.0%) were obtained. There was also varied efficiency in embryo formation (5.44 - 20.96%) with Duma and Chozi wheat varieties showing superiority. Maize variety Kelvedon Glory, had the best response among the maize varieties. Considerable varietal differences in the haploid production efficiency were evident in both wheat and maize varieties. Key Words: Homozygous plants, protocol-efficiency, Triticum aestivum, Zea mays Résumé La technique d’haploide doublé facilite le développement des plantes homozygene dans des limites d’une génération, réduisant extrêmement les temps de reproduction. Bien que le système a été établi avec succès au Kenya, l’amélioration du protocole de rendement reste un critique et facteur limitant potentiellement le succès antérieur. L’objectif de cette étude, ainsi donc, était d’investiguer l’effet du génotype de blé (Triticum aestivum) et du maïs (Zea mays L.) sur le rendement de la production visée d’haploïde en établissant un protocole approprié pour les conditions du Kenya. Vingt génotypes de blé étaient croisés avec six variétés de maïs. Les embryons d’haploide étaient secouris et cultivés pour la regénération de plantes et postérieurement traités avec la colchine pour doublement de chromosomes. Les croix moitié numérotées étaient faites en une cage et le serre et les embryons étaient cultivés dans le laboratoire sous environnement controlé. Les différences génotypiques significatives en graines groupées (51,6-90,0%) étaient obtenues. Il y avait aussi un rendement varié en formation d’embryons (5,44 – 20, 96%) avec les variétés de blé Duma et Chozi montrant une supériorité. La variété de maïs Kelvedon glory, avait la meilleure réponse parmi les variétés de maïs. Des considérables différences variétales dans le rendement de production d’haploides étaient évidentes à la fois dans les variétés de blé et de maïs. Mots Clés: Plantes homozygènes, protocole de rendement, Triticum aestivum, Zea mays Introduction Doubled haploid (DH) system is a biotechnological tool which has been widely applied in wheat breeding programmes (Moieni et al., 1997; Suenaga et al., 1997). The system is developed through haploid production, followed by chromosome doubling, to produce homozygous plants in a single generation. For self-pollinated crops such as bread wheat, a process of genetic fixation for uniformity of agronomic traits after hybridisation is necessary to increase variation. In conventional breeding, six or seven generations are required to achieve homozygosity and genetic stability (Suenaga et al. 1997; Snape, 1998). This long process renders the system inconvenient, and there is need for an alternative process, which takes a shorter time span. Indeed, this has been pointed out by growers in Kenya (Nightingale, 1999 Unpubl.). If well utilised, DH would be a useful breeding tool for meeting this urgent need. The unique genetic property in DH system of allowing homozygous lines to be developed from heterozygous parents, increases the efficiency of a breeding programme by reducing the selection period from 13 to 5 years or less (Suenaga, 1994; Inagaki, 1996). Besides, it provides more accurate and efficient selection of homozygous plants since both dominant and recessive genes are easily expressed in the first generation (Snape, 1998). The new pool of genetic variability created by the DH system is useful in development of new crops. The first criterion for application of DH systems to breeding programmes proposed by Snape et al. (1986) states that DH lines should be produced efficiently from all genotypes. However, the criterion is associated with two problems; the difficulty and efficiency of haploid production through a given DH system, and whether varietal differences in haploid production are negligible. Hence, the need for further investigations to establish varietal efficiency in haploid production Haploids and doubled haploid wheat plants have been repeatedly produced successfully through wheat x maize crosses (embryo rescue system) since the first report by Laurie and Benett (1988). Although this system has been found to be less wheat genotype dependent and more efficient in haploid production than the Hordeum bulbosum system (Laurie and Raymondies, 1991 ; Suenaga et al., 1991), there is a considerable varietal difference in the efficiency among wheat varieties (Inagaki and Tahir, 1990; Laurie and Raymondies, 1991; Suenaga et al., 1991; Suenaga, 1994). It has also been suggested that maize genotypes may affect the efficiency of haploid production (Suenaga and Nakajima, 1989; Suenaga et al., 1991; Ushiyama et al., 1991; Suenaga et al., 1998). In this study, twenty wheat genotypes together with six maize varieties were used for wheat embryo formation and haploid regeneration. Both wheat and maize genotypes are known to influence efficiency in haploid production (Suenaga et al., 1998). Determination and use of high haploid producers will greatly enhance the development of DH lines (Verma et al., 1999). However, the parental effects of these crops on haploid formation are largely unknown. Thus, this study was instituted to investigate the parental effect of wheat and maize varieties/lines on haploid formation and to develop a more efficient DH protocol of wheat using wheat x maize crosses under Kenyan conditions. Materials and methods Twenty bread wheat (Triticum aestivum) genotypes (Table 1) and one F1 hybrid maize Kelvedon Glory (Zea mays L.) were used as female and pollen parent, respectively, to analyse varietal differences of wheat in embryo formation. The pedigree and source of these genotypes are as indicated in Table 1. The wheat genotypes were planted in pots in the greenhouse under temperatures of day/night 22-25°C/ 10-17°C with 16-hour day length. Each variety/line had two pots with an average of 9 plants per pot. The spikes for each genotype were used as replicates for analysis. Six maize varieties (Table 2) were used as pollen parents to analyse the effect of the maize genotype on the formation of wheat embryos. Two months before wheat planting, maize materials were grown in the field and variety Chozi was used as female parent. Half-diallel crosses were made between wheat and maize varieties. Wheat spikes were emasculated 2-3 days before pollination and covered with glassine papers. During the emasculation the central floret was removed, awns excised and glumes were pruned. Hand-pollination was done with freshly harvested pollen, while the stigmas were receptive. The spikes were subsequently covered with polythene bags after pollination and 2,4-D (100 mg l-1) was injected into the uppermost internodes of the wheat stems a day after pollination. Polythene papers were replaced with glassine papers five days later as described by Kinyua et al. (2000). From the 16th to 18th day after pollination, embryo development was investigated microscopically. An average of five spikes for each genotype was harvested. Number of florets pollinated, seeds developed and embryos rescued were recorded. Seeds were sterilised using 70% ethanol for 2 minutes and 20% Clorox for 8 minutes. Embryos were aseptically excised and transferred into culture vessels containing 10 ml of enriched MS medium (Murashige and Skoog, 1962) modified with filter sterilised B5 vitamins (Kinyua et al., 2000). Sucrose 2% as the carbon source and phytagel (3 g) for gelling agent were added and cultures were stored in the refrigerator at 4°C for 5 days to enhance embryo germination before incubation at 25°C until germination (5-7 days). After germination, embryos were transferred to the growth chamber with controlled temperatures of 25°C until the 2-leafed stage (about 3 weeks). Seedlings with normal roots and shoots were selected and transplanted in sterilised potted soil under plastic cover where they were maintained for 3 days. They were then transferred to the greenhouse (22-25°C). At tillering stage chromosome doubling of haploid plants was stimulated by treating the plantlets with colchicine for five hours (Inagaki, 1986) after confirming the ploidy level of the plants and rinsing with water for 3 hours. Subsequently, plants were left to grow to maturity. The same procedure was repeated for maize genotypes. Percentage seed set (seeds developed / florets pollinated) x 100), percentage embryos developed (obtained/seeds developed) x 100) and efficiency of embryo formation (embryos obtained / florets pollinated) x 100) were calculated. These data were arcsine transformed before analysis of variance to detect differences in efficiency between the wheat genotypes and the maize genotypes. Results Wheat genotype effect. Table 3 shows the effect of wheat genotype on seed set (%), embryo development (%) and efficiency of embryo development (%) for each variety. All the investigated varieties were crossable and, hence produced embryos. Embryo formation, however, ranged from 1.59 to 31.27%. Varietal differences for embryo development were highly significant among the varieties (P<0.010), while there were no significant differences in the percentage of seed set. Seed set and embryo formation were not significantly correlated (P=0.05) (Table 4). The overall efficiency of embryo formation varied from 5.44% for K7872 to 20.79% for Chozi both varieties from Kenya. Variety Duma and Chozi showed superiority in embryo development. Maize genotype. The effects of maize genotype on the formation of wheat embryos are given in Table 5. For embryo development, the percentages ranged from 9.1% for F1 Hybrid yellow to 23.58% for F1 Hybrid Kelvedon Glory. A significant large varietal difference (P<0.026) was observed among maize varieties. Overall efficiency ranged from 8.91-19.28%. No significant difference was noted for the seed set. Effect of F1 lines on embryo formation. A comparison was made on F1 and their parents (Table 6) and the results are in agreement with earlier observations reported by Suenaga (1994) that the F1 progeny show intermediate embryo formation compared to the parents. However, out of the four lines evaluated, only R810/Kongoni exhibited some heterosis effects. Discussion For large scale DH production, efficiency of haploid production is of paramount importance. Suenaga et al. (1998) stated that one of the ways of increasing DH production efficiency is to improve the percentage of haploids per floret. In order to increase the efficiency of wheat x maize crosses, a range of local and elite wheat and maize varieties was evaluated. Thus, this study revealed that percentage seed set and embryo formation were significantly different among wheat and maize varieties. These results are consistent with those reported by others (Inagaki and Tahir, 1990; Laurie and Reymondies, 1991; Suenaga, 1994; Suenaga, 1998) who detected a considerable genotype effects. Among the varieties and lines evaluated, the drought tolerant varieties Chozi (23.58%), Duma (22.46%) and Ngamia (18.53%) responded better to haploid embryo development than the conventional wheat varieties. The results indicate that the three varieties could be used in a breeding programme to improve the efficiency of haploid production. For the maize varieties tested, there was no significant difference among the local varieties but there was a considerable difference in comparison to Kelvedon Glory F1 an Australian maize variety. These results indicate that there is need to evaluate a wider range of maize cultivars to detect one with higher efficiency or improve the local maize with elite germplasm for haploid efficiency. Preliminary work done on F1 crosses of four haploid lines indicated that there could be heterosis effects on embryo formation after crossing. Hence, there is need for further work to study haploid production efficiency of developed lines after hybridisation. References
©2002, African Crop Science Society The following images related to this document are available:Photo images[cs02029t2.jpg] [cs02029t1.jpg] [cs02029t5.jpg] [cs02029t4.jpg] [cs02029t3.jpg] [cs02029t6.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}