 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 11. No. 2, 2003, pp. 87-95 GENETIC VARIATION IN BANANA CULTIVAR 'SUKALI NDIZI' GROWN IN DIFFERENT REGIONS OF UGANDA M. Pillay, C. Dimkpa, G. Ude1, D. Makumbi and W. Tushemereirwe2 International Institute of Tropical Agriculture, Eastern and Southern Africa
Regional Center,
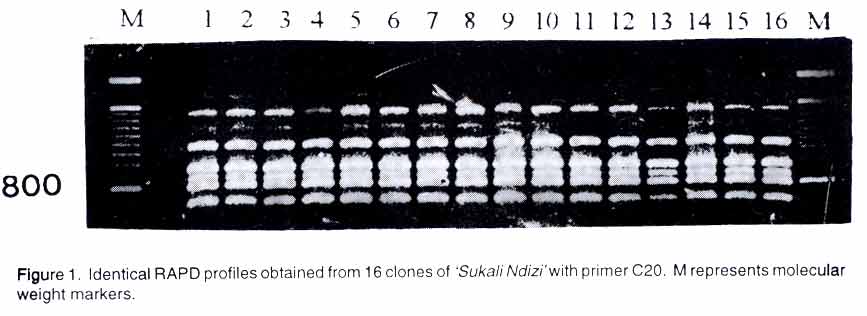
P. O. Box 7878, Kampala, Uganda (Received 13 May, 2002; accepted 19 March, 2003) Code Number: cs03011 ABSTRACT Banana is an important food and cash crop in Uganda. The crop displays wide diversity but the genetic relationship between and withing cultivars is largely unknown. A study was conducted to assess genetic relationships among 'Sukali Ndizi' clones collected from 16 different localities in Uganda using the random amplified polymorphic DNA (RAPD) and amplified fragment length polymorphism (AFLP) techniques. Thirty-four RAPD primers used singly and in combination produced 234 unambiguous bands. The RAPD primers produced identical banding patterns in all the samples and were, therefore, not useful in differentiating the clones. However, the 9 AFLP primer pairs produced 554 fragments, 17 of which were polymorphic. Genetic relationships were established from the AFLP data by cluster analysis. Two groups of clones were clearly defined. Clones from selected contiguous districts such as Lira and Soroti, Bushenyi and Kasese districts were highly similar. Other closely related clones were from disjunctive localities. The similarity of the clones in adjacent districts is attributed to local exchange of germplasm, while similarity of clones in non-contiguous districts is probably the result of more purposeful and selective transfer of planting material. In general, the observed clonal variation may be due to somatic mutations. Key Words: AFLP, germplasm, Musa, primers, RAPD, somatic mutations, Uganda RÉSUMÉ La banane est un important aliment et plante commerciale en Ouganda. La plante affiche une large diversité mais la relation génétique entre et avec les variétés est largement inconnue. Une étude était conduite pour évaluer les relations génétiques entre les clones «Sukali Ndizi» collectés dans 16 localités différentes de l'Ouganda en utilisant un prélévement aléatoire polymorphique amplifié DNA (RAPD) et des techniques amplifiés de polymorphisme de longueur de fragment (AFLP). Trente quatre premiers RAPD utilisés séparement et en combinaison ont produit 234 bandes non ambigues. Les premiers RAPD ont produit des tendances identiques en bandes dans tous les échantillons et étaient, ainsi non utiles dans la differentiation des clones. Cependant, les premières paires d'AFLP par l'analyse de bouquet. Deux groupes des clones étaient clairement définis. Les clones des districts contigus sélectionnés tels que Lira et Soroti, Bushenyi et Kasese étaient fortement similaires. Les autres clones fortement apparentés étaient des localités disjointes. La similarité des clones des districts adjacents est attribué à l'échange local des germoplasme, pendant que la similarité des clones dans les districts non-contigu est probablement le résultat d'un transfert plus objectif et plus selectif du matériel de plantation. En général, la variation clonale observée peut être due aux mutations somatiques. Mots Clés: AFLP, germoplasme, Musa, premiers, RAPD, mutations somatiques, Ouganda INTRODUCTION Banana (Musa sp.) is an important food and cash crop in Uganda. The East African highlands are considered a secondary centre of diversity for the AAA cooking bananas, known locally as 'matooke' (Swennen et al., 1995). In addition to these endemic bananas, a number of exotic or introduced cultivars are present in the region and are sold primarily as dessert bananas, although some growers have pure stands for the sole purpose of beer production. One example of such a cultivar is 'Sukali Ndizi' that is characterized by its slender, green-yellow stems and small short and plump fruits (Karamura and Karamura, 1994). The name Sukali is associated with 'sukari' in Arabic and Swahili that means sugar because of the sweetness of the ripened fruit of this cultivar. 'Sukali Ndizi' and other similar cultivars were introduced into Uganda in the 1900s (Mukasa, 1970). The Botanical garden at Entebbe could have played a role in the introduction and diffusion of these exotic banana cultivars in Uganda (Rossel, 1998). 'Sukali Ndizi' has been grouped with other AB cultivars like 'Kisubi' and is classified as belonging to the 'Ney poovan' group (Stover and Simmonds, 1987). Like other bananas in the region, it is affected by a number of pests and diseases including black Sigatoka (Mycosphaerella fijiensis), Fusarium wilt (Fusarium oxysporum), a complex of nematodes (Radopholus similis, Helicotylenchus multicinctus, and Pratylenchus goodeyi), weevils (Cosmopolites sordidus) and banana streak virus (De Vries and Toenniessen, 2001). An understanding of the amount and distribution of the genetic variation within the genepool of 'Sukali Ndizi' will be valuable for the success of breeding for improved cultivars. Molecular markers including random amplified polymorphic DNA (RAPD) and amplified fragment length polymorphism (AFLP) have been used to assess genetic variation in a number of crop species. The RAPD technique involves the amplification of DNA segments using random sequence primers, generally of ten bases, to find polymorphic regions within the genome defined by the primer sequence (Williams et al., 1990). The objectives of this study were to use RAPD and AFLP markers to: examine DNA variation in 'Sukali Ndizi' clones, and demonstrate genetic relationships among the clones. MATERIALS AND METHODS Plant material. The plant material consisted of clones of 'Sukali Ndizi' collected from 16 geographical localities in Uganda (Table 1) and kept among the germplasm collections of Kawanda Agricultural Research Institute in Uganda.
Genomic DNA isolation and RAPD analysis. Isolation of genomic DNA was carried out as described by Crouch et al. (1998) and RAPD analyses were done following the method in Pillay et al. (2001). Thirty-four single primers and 8 combinations of 2 primers each were used to amplify the DNA samples (Table 2).
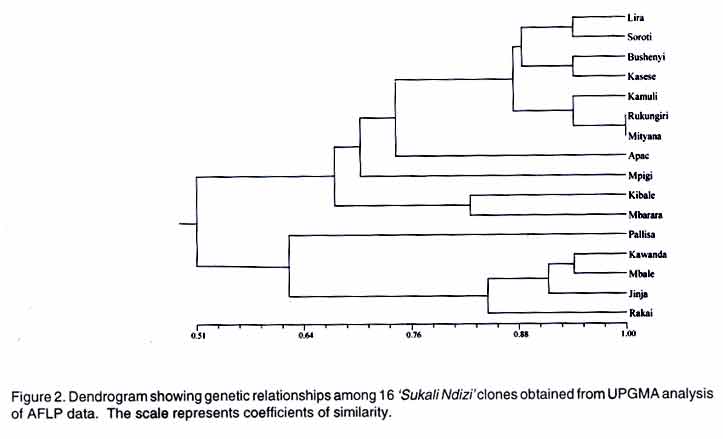
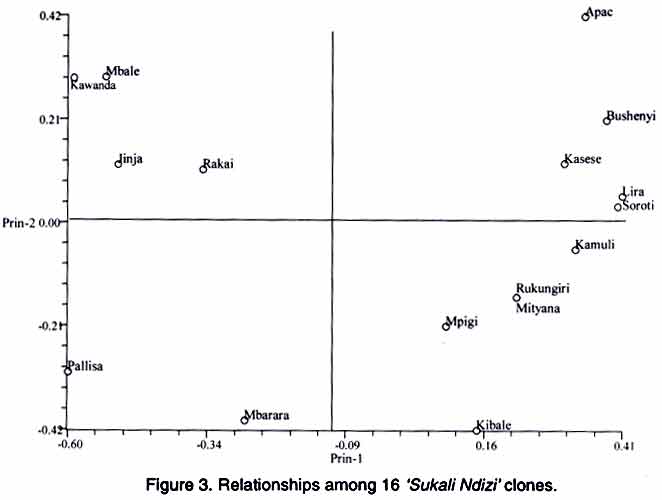
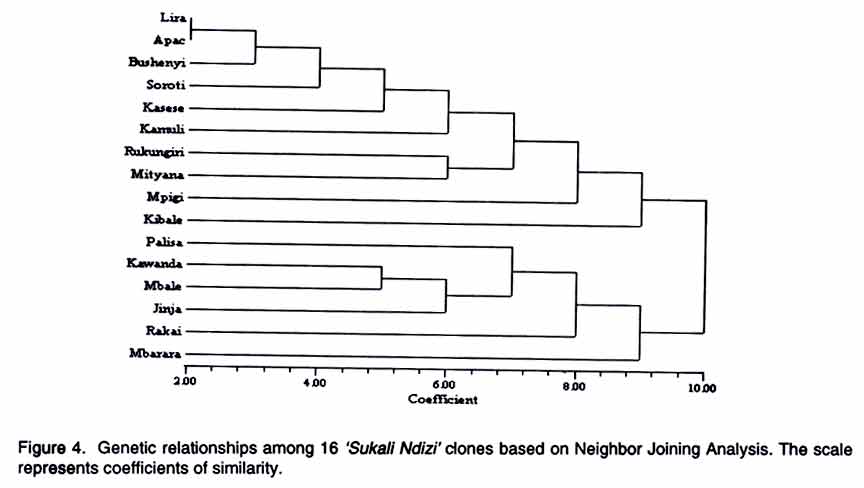
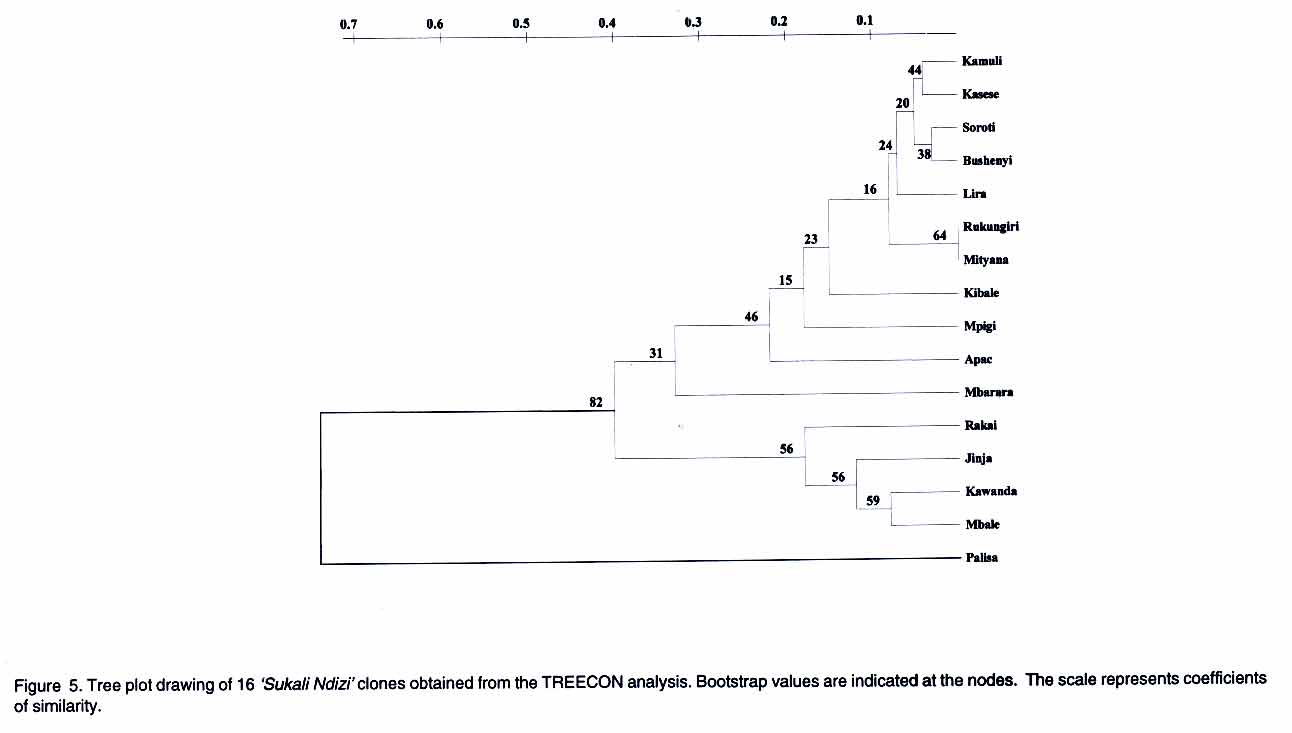
AFLP analysis. AFLP analysis was carried out as described by Ude et al., (2002). The following selective bases from the GIBCO BRL kit (Life Technologies, Paisley, Scotland) for AFLP EcoRI plus 3 (E) and MseI plus 3 primers (M), respectively, were used: E-AGC/M-CTC, E-ACC/M-CAC, E-ACT/M-CAT, E-ACA/M-CAA, E-AGC/M-CAA, E-AAC/M-CAG, E-ACC/M-CTT, E-AGC/M-CTT and E-ACA/M-CTG Data analysis. Polymorphic fragments in the AFLP gels were scored as present (1) or absent (0) to produce a binary data matrix. A fragment was considered polymorphic if present in at least one clone and absent in the others. Genetic similarity values between pairs of clones were calculated using the simple matching coefficients from the NTSYS-pc software package (Rohlf, 1994). The genetic similarity matrix was used to generate a dendrogram using the unweighted pair-group method of arithmetic averages (UPGMA) and the Neighbour Joining (NJ) method. The data were also subjected to Principal Co-ordinate Analysis. Since the NTSYS-pc package does not provide bootstrap analysis, the data were also analysed with TREECON, a software package for the construction and drawing of evolutionary trees (Van de Peer and De Wachter, 1994). RESULTS Rapid amplified polymorphic DNA (RAPD). The 34 single primers and 7 combinations of 2 primers each amplified a total of 234 clearly defined fragments ranging in size from 300 bp to 2.07 kbp. No DNA polymorphisms were detected among the 16 clones. All the amplification products were monomorphic. A representative RAPD profile is shown in Figure 1. Amplified fragment length polymorphism (AFLP). The nine primer pairs produced 554 fragments, ranging in size from 50 to 500 bp. Only 17 of these fragments were polymorphic. The dendrogram derived from UPGMA divided the 16 geographical clones into two groups (Fig. 2). Group 1 was composed of clones from Lira, Soroti, Bushenyi, Kasese, Kamuli, Rukungiri, Mityana, Apac, Mpigi, Kibale and Mbarara districts. Group 2 consisted of clones from Pallisa, Kawanda, Mbale, Jinja and Rakai districts. The clones from Lira, Soroti, Bushenyi, Kasese, Kamuli, Rukungiri and Mityana formed a cluster within Group 1. This is illustrated more clearly in the scatter-plot diagram (Fig. 3). Within group one, sister-group relationships existed between the clones from (a) Lira and Soroti, (b) Bushenyi and Kasese, (c) Kamuli, Rukungiri and Mityana and, (d) Kibale and Mbarara. Clones from Rukungiri and Mityana produced identical fragment patterns with all the primer pairs and could not be differentiated (Figs. 2 and 3). In group two, the clones from Kawanda and Mbale appeared closer, while the clone from Pallisa was the most distinct. The close grouping of the clones from Kawanda, Mbale, Jinja and Rakai and the distinctness of the clone from Pallisa is evident in Figure 3. The NJ analysis also clustered the clones into the two large groups with one exception. The clone from Mbarara was hence included in group 2 (Fig. 4). The NJ analysis also grouped clones from Lira and Apac together, whereas these clones appeared more distant in the UPGMA analysis. Other differences in the NJ analysis are presented in Figure 4. The tree generated with the TREECON computer programme is shown in Figure 5. Two major cluster of clones were also evident. With the exception of the clone from Pallisa that appeared as an outgroup, the composition of the two main clusters was similar. The sister group relationships portrayed by the TREECON analysis were different from those in UPGMA (Figs. 2 and 5). For purposes of discussion, only the tree derived from UPGMA is presented. The cophenetic correlation was r = 0.81. DISCUSSION Although RAPD markers have been widely used to detect polymorphisms in a large number of crop species (Kaemmer et al., 1992; Bhat and Jarret, 1995; Pillay et al., 2001; Ude et al., 2001), this is the first study in which RAPD markers have been used to detect DNA variation among clones of a banana cultivar. It appears that the RAPD technique is not efficient for detecting clonal variation in banana. One of the advantages of the RAPD method is that the arbitrarily designed primers can potentially anneal to homologous sequences in the entire genome, providing greater opportunities to uncover polymorphic regions (Williams et al., 1990). The RAPD markers are considered to be more prevalent in repeated DNA sequences and, therefore, in non-coding regions which are more susceptible to mutations (Irwin et al., 1998). If the latter assumption is correct, one would have expected to find polymorphisms among 'Sukali Ndizi' clones. Inability to detect DNA variation is, perhaps, not due to the RAPD technique itself but to the limited number of primers used in this study. On the contrary, AFLP technique was useful in detecting genetic variation in this closely related material. The similarity of the clones in adjacent districts could be attributed to local exchange of germplasm while more purposeful and selective transfer of planting material may explain similarity of clones in non-contiguous districts. Unlike RAPD, the AFLP fingerprinting technique detects genomic restriction fragments that result from the simultaneous digestion of the DNA with two restriction enzymes. In this study, 17 polymorphisms were detected out of 554 amplified restriction fragments after amplification with 9 primer pairs. Assuming that a single clone of 'Sukali Ndizi' was introduced in Uganda, then the observed DNA variation among the 16 clones collected from the different localities is likely a result of somatic mutations. The alternative explanation is that a number of genetically different clones was initially introduced into Uganda and the DNA variation among the 16 clones reflects these differences. Our study provides a rough estimate of mutation frequencies in banana. The ratio of the number of polymorphisms to total number of fragments in this study is 0.03. If 'Sukali Ndizi' was introduced into Uganda in early 1900, the plant would have passed through approximately 100 propagation cycles giving a mutation frequency, F = 0.0003. Diversity of bananas in Africa is considered to be the result of accumulated somatic mutations (Simmonds, 1966). Although the exact mechanisms of these mutations remain unknown, it is probable that transposable elements may be involved. Retrotransposons have been identified in banana (Balint-Kurti et al., 1999; Khayat et al., 1999). Involvement of retrotransposons in spontaneous mutations has been reported for maize (Varagona et al., 1992) and rice (Hirochika, 1997). It is also known that retrotransposons are activated by stress and environmental factors (Grandbastien, 1998). Karamura (1998) reported that the contrasting physiogeography and climates of East Africa and the social backgrounds of the region might have played a role in the diversification of banana clones in the region. This may be true also for 'Sukali Ndizi'. Diversity within a genotype may be a safeguard against the outbreaks of epidemics. Spontaneous mutations occurring in plants and the concomitant genomic changes broaden the genetic base and are a vanguard against vulnerability to attacks by pests and diseases. However, it appears that such genetic changes in 'Sukali Ndizi' have had no influence on the plant's phenotype. A close examination of the plants used in this study showed no apparent morphological variation. This study has important implications for germplasm conservation and establishment of core collections. A core collection consists of a limited set of accessions derived from an existing germplasm collection, chosen to represent the genetic spectrum of the whole collection (Brown, 1995). A core collection should represent the genetic diversity of a crop species. Our samples showed no distinct phenotypic variation and would appear to be identical genetically, yet they were quite different at the molecular level. Since they are phenotypically similar, a plant collector would select any one of these clones as an entry for the core collection. However, our study showed that one clone would not represent the spectrum of molecular diversity identified in this cultivar. Molecular characterization is essential to ascertain this variation. Therefore, future studies on Musa should not only collect plants on the basis of morphology alone but consideration should be given to the 'hidden' variation. In conclusion, our study provides a basis for banana breeders to select parents of 'Sukali Ndizi' on the basis of genetic diversity. Further studies using additional accessions from a broader geographical distribution would provide valuable information on the available genetic diversity in this cultivar. From this study, the AFLP technique is more useful than RAPD in detecting variation in closely related material in banana. ACKNOWLEDGEMENTS Funding for this research was provided, in part, by a grant from the Directorate General for International Cooperation (DGIC), Belgium. REFERENCES
©2003, African Crop Science Society The following images related to this document are available:Photo images[cs03011f5.jpg] [cs03011f4.jpg] [cs03011f1.jpg] [cs03011f2.jpg] [cs03011f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}