 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
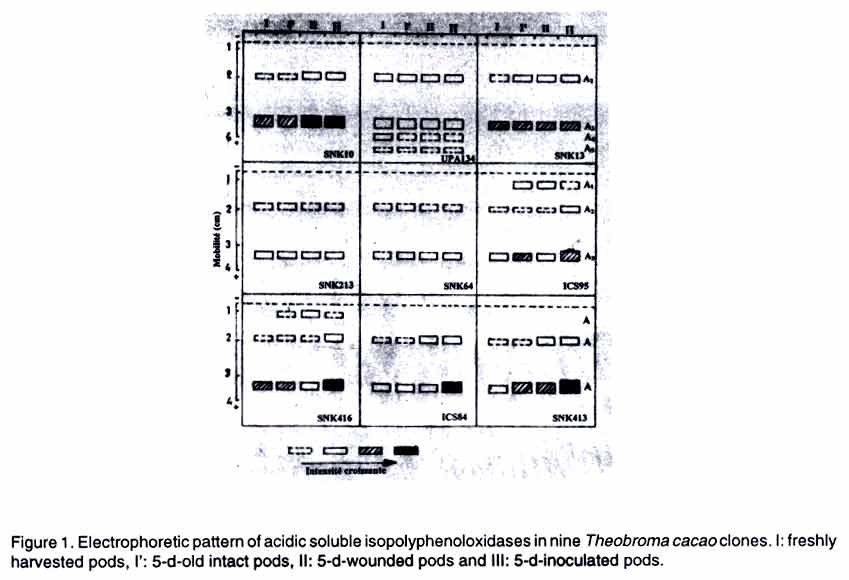
African Crop Science Journal, Vol. 11. No. 2, 2003, pp. 97-106 VARIATION OF b-1,3-GLUCANASE, CHITINASE AND POLYPHENOLOXIDASE ACTIVITIES IN CACAO PODS UPON Phytophthora megakarya INOCULATION N. D. Omokolo, D. J. Nankeu, N. Niemenak and T. Boudjeko Higher Teacher Training College, University of Yaoundé I, P.O. Box 47, Yaoundé, Cameroon (Received 15 March, 2002; accepted 13 March, 2003) Code Number: cs03012 ABSTRACT Cacao (Theobroma cacao L.) clones that differ in susceptibility to black pod disease were analysed for response to stress induced by pod inoculation with the fungus Phytophthora megakarya Braz. Et Griff. Fungal inoculation significantly stimulated b-1,3-glucanase activity in both soluble and ionically-bound fractions of the less susceptible clones. No significant correlation was found in the other clones. Chitinase activity in both soluble and ionically-bound fractions was generally reduced upon pod inoculation with the fungus, but this pattern did not correlate with disease severity. Polyphenoloxidase activity, following pod inoculation, was genotype dependent in the soluble fractions, whereas in the ionically-bound fractions, stimulation of this activity was recorded in the less susceptible clones. Moreover, pod inoculation was characterised by the appearance of a new polyphenoloxidase isoform A'2 in the less susceptible clones. Key Words: Absorbance, cortex, ionically-bound, Theobroma cacao, wonding RÉSUMÉ Les réponses aux stress induits par innoculation de gousses de cacao (Theobroma cacao L.), aux susceptibilités differentes à la maladie de Blackpod, par le Phytophthora megakarya Braz étaient analysées. L'inoculation de fungi a significativement stimulé les activités de « glucanase » b-1,3 dans les deux fractions solubles et à liaison ionique des clones moins susceptibles. Pas des corrélations significatives étaient observées dans d'autres clones. L'activité de la chitanase dans les deux milieux était generalement reduite juste après l'inoculation par le fongi, mais cette tendance n'a pas corrélée avec la sévérité de la maladie. L'activité du polyphenoloxidase, après inoculation de gousses, était généralement dependante du génotype dans la fraction soluble, alors que la stimulation de cette activité était observée dans les clones moins susceptibles. En plus, l'inoculation des gousses était caracterisée par l'apparition de nouveau polyphenolosidase isoformes A'2 dans les clones les moins susceptibles. Mots Clés: Absorbance, cortex, ionically-bound, Theobroma cacao, wonding INTRODUCTION Cacao (Theobroma cacao L.) is an important commercial crop in Cameroon (Anonyme, 2000). Cacao production is constrained by several disease infestations which are estimated to cause losses as high as 80% in Cameroon (Berry and Cilas, 1994) and 100 % in Ghana (Dakwa, 1988).However the epidemeology of these diseases is largely unknown. For instance, many plant breeders have addressed the highly destructive cocoa black pod caused by Phytophthora megakarya, but a lot of questions concerning the mechanisms of disease tolerance are yet to be elucidated. The ubiquitous plant response to the pathogen is the activation of some physiological and biochemical processes necessary for impeding further pathogen progress. These include alteration of ion fluxes across the plant cell membrane, generation of active oxygen species, change in the phosphorylation state of regulatory proteins, and transcriptional activation of plant defence system. These reactions culminate in local accumulation of phytoalexins and deposition of structural polymers such as callose, lignin and suberin, as well as cell death at the site of infection (Benhamou, 1995; Benhamou and Nicole, 1999). In addition, various novel proteins known as pathogenesis related proteins (PRs) are induced (van Loon et al., 1994). Some of these proteins have been identified as b-1,3-glucanases (EC 3. 2.1.39) and chitinases (EC 3.2.1.14). It has been reported that in some species, following infection, resistant genotypes accumulate these enzymes more rapidly than susceptible genotypes (Irving and Kuc, 1990; Schnöder et al., 1992). It is postulated that b-1,3-glucanases participate in the active defence response of plants to pathogens, either as direct antimicrobial molecules in the break-down of pathogen cell wall (Keen and Yoshikawa, 1983; Mauch et al., 1988), or indirectly by releasing elicitors from fungal cell wall (Boller, 1987,1988). Chitinases degrade chitin, a homopolymer consisting of b-1,4-N-acetylglucosamine units and which is found in virus capsides, most bacteria and fungi (Metraux and Boller, 1986). In vitro growth of a number of fungi can be delayed by the addition of glucanase and chitinase in the culture media (Arlorio et al., 1992). Another group of defence proteins include oxidative enzymes such as polyphenoloxidases (PPO), copper enzymes that catalyse, through the reduction of molecular O2, the hydroxylation of monophenols to O-diphenols (cresolase activity, E.C.1.14.18.1) as well as the oxidation of O-diphenols to O-quinones (catecholase activity, E.C.1.10.3.2). The generated oxidized phenols contribute to the formation of defence barriers against pathogens (Mayer and Harel, 1979). Although these proteins have been well investigated in herbaceous species, they have rarely been studied in woody plants. Their role in the defence mechanism in woody plants was demonstrated in grapevine after infection by Botrytis cinerea (Renault et al., 1996). There is a paucity of information about the quantitative and qualitative levels of these enzymes during black pod disease development, caused by P. megakarya on cocoa (Theobroma cacao). Thus, the objective of this study was to investigate the variation of b-1,3-glucanase and chitinase activities as well as characterise isopoly-phenoloxidases in the cortex of nine clones of T. cacao, after pod inoculation with P. megakarya MATERIALS AND METHODS Cocoa and fungus materials. Nine cocoa clones used in this study were selected from a field plot at Institut de Recherche Agricole pour le Développement (IRAD ) at Nkolbisson, Yaounde, Cameroon. These clones differ in susceptibility to black pod disease (Blaha and Lotode, 1976), where the susceptibility is expressed by the rate of increase in the necrotic area following pod infection. The clones were: SNK10, UPA134, SNK13 (highly susceptible); SNK213, SNK64, ICS95 (mildly susceptible) and SNK416, ICS84, SNK413 (less susceptible). Three-month-old pods were used. The isolate of P. megakarya used was the strain L2C2 isolated from the Yaounde area and obtained from the microbiology laboratory at IRAD. The fungus was routinely cultured in a medium containing 31.8% agar, 20%V8 juice and 0.3% CaCO3, to maintain its virulence. Inoculation. Cocoa pods were washed thoroughly with-tap water and blotted dry with Whatman No. 3 paper. They were then surface sterilized with ethanol 70°. For each clone, pods were divided into three lots of ten pods each. Lot I with intact pods, Lot II with pods inoculated with sterile agar (wounded), and Lot III with pods inoculated with fungal cultures on agar (infected). Agar disks (6 mm diameter), cut from 7-day-old fungal cultures, were inserted into holes (0.7 cm in diameter) cut in the pod with a cork borer on two opposite points located at the distal part of the fruits. The holes, containing agar disks, were covered with wet cotton. Pods were then wrapped with aluminium foil and left at room temperature for 5 days, since it has been demonstrated that significant effects appear at day 5 after pod inoculation (Nyassé et al., 1995). Preparation of crude enzyme extracts. At days 0 and 5 after inoculation, cortex samples were collected at 1 cm beyond the necrosis, for biochemical analyses. Samples from pods of the same clone and of the same treatment, were combined and stored at -20°C until analysis. For extraction of soluble b-1,3-glucanase and chitinase, 3 g of frozen tissues were ground in acetone, in a pre-chilled mortar, at 4°C and filtered under a reduced pressure. The resulting powder was homogenised in 3 ml of 50 mM sodium acetate buffer pH 5.2 containing 0.5 M mannitol and 10 mM b-mercaptoethanol. The homogenate was centrifuged (centrifuge HERMLE ZK510) at 10,000 x g for 20 min. The supernatant was collected and the pellet resuspended in 20 mM sodium acetate buffer pH 5.2 containing 1 M NaCl and 10 mM b-mercaptoethanol for extraction of ionically-bound b-1,3-glucanases and chitinases. The homogenate was incubated at 4 °C for 1 h and centrifuged at 10,000 x g for 20 min. To remove carbohydrates which generally interfere with b-1,3-glucanase and chitinase measurements, the two supernatants were brought to 95% saturation with (NH4)2SO4 at 0 °C for 2 h, according to Mauch et al. (1988). The resulting precipitate was collected by centrifugation at 10,000 x g for 10 min and suspended in 50 mM sodium acetate buffer pH 5.2. Soluble and ionically-bound enzyme extracts were dialyzed twice against 20 mM sodium acetate buffer pH 5.2 at 4°C for 14 h. The dialysed extracts were used for b-1,3-glucanase and chitinase assays. To extract soluble polyphenoloxidases, 50 mM Tris-maleate buffer pH 7 containing 0.5 M mannitol was added to the original powder in the proportion 3:1 (v:w), stirred for 2 min and incubated at 4°C for 20 min. After centrifugation at 10,000 x g for 20 min, the supernatant containing soluble PPO was collected and kept at -20 °C until measurements. Ionically-bound PPO fraction was obtained by mixing the residue in 10 mM Tris-maleate buffer pH 7 containing 1 M NaCl. The homogenate was incubated at 4°C for 1 h and then centrifuged at 10,000 x g for 20 min. Enzyme assay and protein determination. Enzyme activities were analysed in both soluble and ionically-bound fractions since their relative activities depend on the physiological state of the cells. The b-1,3-glucanase activity was determined by measuring the rate of release of reducing sugars using laminarin as substrate and glucose as standard. The reaction mixture consisted of 800 µl of 20 mM sodium acetate buffer pH 5.2, 100 µl laminarin 1% and 100 µl of the dialyzed enzyme extract. After 1 h of incubation at 50°C, total reducing sugars were assayed by the colorimetric method of Somogyi (1952). A unit of enzymatic activity is the amount of enzyme releasing 1 mole of glucose equivalent min-1. Chitinase activity was assayed by measuring the rate of N-acetylglucosamine production using chitin as substrate. The reaction mixture consisted of 850 µl of 20 mM sodium acetate buffer pH 5.2, 100 µl of chitin prepared by the method of Rupley (1964) and 50 µl of dialysed enzyme extract. After 1 h of incubation at 50°C, the release of N-acetylglucosamine was estimated by the colorimetric method of Somogyi (1952). A unit of enzymatic activity is the amount of enzyme releasing 1 mole N-acetylglucosamine min-1. Assay of PPO was carried out using catechol as substrate. The standard assay contained 50 µl of the enzyme extract and 3 ml of 0.66 M phosphate buffer pH 7. The mixture was transferred quickly in a cuvette after adding 0.3 ml catechol 10 mM and the increase in absorbance at 330 nm over a 30 sec. period was determined using a spectro-photometer (U-2000 Hitachi). Enzyme activity is expressed as change in absorbance: DAbsorbance 30 s-1 g-1 fresh weight. Proteins were estimated according to Bradford (1976). All the concentrations indicated are final concentrations in assay media. PAGE gel staining. Isopolyphenoloxidases were analysed by PAGE under non-reducing conditions. Acrylamide (12 and 5%) was used for the resolving and stacking gel, respectively. Basic gel buffer was 0.4 M Tris pH 8.8 and the electrode buffer 0.03 M Tris, 0.2 M glycine pH 8.6. Samples contained about 20 µg proteins and 10% (v:v) glycerol. The gel was run at 4 ± 1°C at a constant voltage (100 V) for 19 h and then stained in 100 mM sodium acetate buffer pH 5, 10 mM catechol and 5 mM DL DOPA for 30 min. Data analysis. The study was conducted twice and the treatments replicated thrice each time. The data collected were analysed by the analysis of variance (ANOVA). RESULTS b-1,3-glucanase activity. Soluble b-1,3-glucanase activity was higher in the freshly harvested pods of clones ICS84 and SNK413 than in the 7 other clones (Table 1). This activity ranged from 0.879 units 10-4g-1 fresh weight (clone SNK416) to 1.573 units 10-4g-1 fresh weight (clone SNK413). In 5-day-old intact pods, changes due to aging were genotype-dependent. Glucanase activity increased in the pods of SNK10 (70%) and SNK13 (24%); decreased in the clones SNK64 (59%), ICS95 (31%), SNK416 (22%), ICS84 (72%) and SNK413 (65%), and remained almost unchanged in UPA134 and SNK213 pods. When 5-day-old intact pods were compared with 5-day-old wounded pods, the activity of soluble glucanase increased in clones ICS84 (52%) and SNK413 (30%), decreased in the clones SNK10 (24%), UPA134 (67%) and SNK13 (23%), while remaining almost constant in clones SNK213, SNK64 and ICS95.
Means with same letter in the same line are not significantly different at 5 % level (Each value is mean +/- S.E. of three replicates) Analysis of variance between 5-day-old intact pods and 5-day-old inoculated pods showed a statistical difference in the activity of soluble b-1,3-glucanase for all the clones studied except for clones SNK64 and ICS95. This activity increased in clones ICS84 and SNK413 but decreased in the clones SNK10, UPA134, SNK13, SNK213 and SNK416. When the activity of b-1,3-glucanase in 5-day-old wounded pods were compared with that in 5-day-old inoculated pods, statistical differences were observed in clones SNK10, SNK13, SNK213, SNK416 (where it decreased) and in clones ICS84 and SNK413 (where it increased). The activity of soluble b-1,3-glucanase in inoculated pods was higher in the clone SNK413 than in the eight other clones. Clone UPA134 displayed the lowest activity (Table 1). The activity of ionically-bound b-1,3-glucanase ranged from 0.142 units 10-4 g-1 fresh weight (SNK416) to 0.252 units 10-4 g-1 fresh weight (ICS95) in freshly harvested pods (Table 2). During pod aging, this activity remained unchanged in all the clones, except for clones UPA134, ICS84 and SNK413 where it decreased. When 5-day-old intact pods were compared with 5-day-old wounded pods, the ionically-bound b-1,3-glucanase activity decreased in clones SNK213 (29%), SNK64 (22%) and ICS95 (41%), yet it remained almost constant in the six other clones (Table 2).
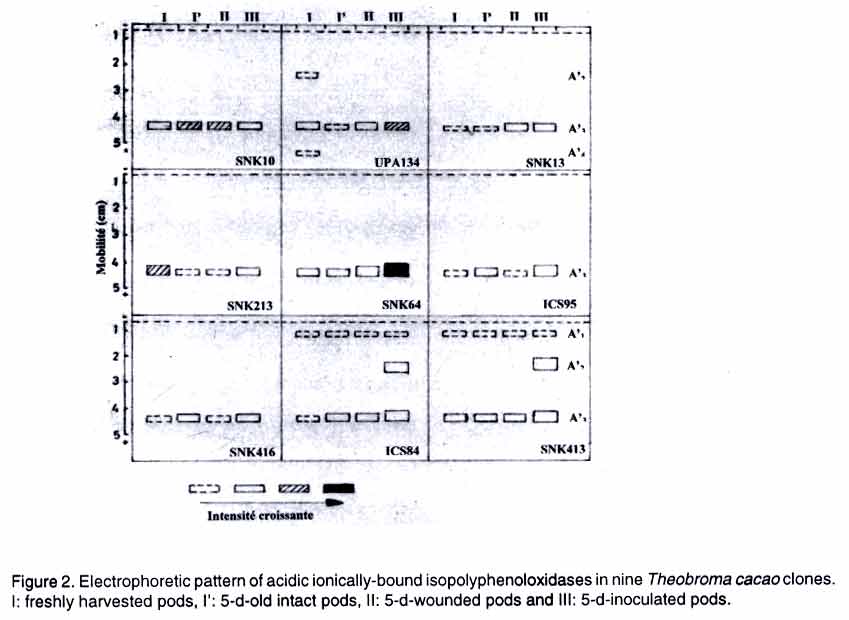
Means with same letter in the same line are not significantly different at 5 % level (Each value is mean +/- S.E. of three replicates) Between 5-day-old intact pods and 5-day-old inoculated pods, statistically significant decreases in the activity of ionically-bound b-1,3-glucanase were observed in the clones SNK10, SNK213, SNK64 and ICS95, while increases were recorded in the clones ICS84 and SNK413. The last two clones also displayed an increase in activity when 5-day-old wounded pods were compared with 5-day-old inoculated pods, but a decrease was recorded in the clone SNK64. No significant differences were found in the other clones. Among the inoculated pods, ICS84 and SNK413 pods displayed the highest ionically-bound b-1,3-glucanase activity (0.209 and 0.194 units 10-4 g-1 fresh weight, respectively). Clone SNK64 had the lowest activity (0.108 unit 10-4 g-1 fresh weight) (Table 2). Chitinase activity. Soluble chitinase activity in freshly harvested pods was highest in the clones SNK64 and SNK413 (Table 1). The activity ranged from 0.557 units 10-4 g-1 fresh weight (SNK416) to 1.000 units 10-4 g-1 fresh weight (SNK64). Chitinase activity in intact pods increased after five days in clones SNK10 (46%) and SNK13 (22%), but decreased in pods of SNK64 (55%), ICS84 (61%) and SNK413 (45%). Pod aging did not affect pods activity of UPA134, SNK213, ICS95 and SNK416. Compared to that of 5-day-old intact pods, the soluble chitinase activity of 5-day-old wounded pods decreased in clones UPA134 (22%) and SNK13 (35%). Soluble chitinase showed no significant activity change in the seven other clones after wounding (Table 1). In 5-day-old intact and 5-day-old inoculated pods, there was a high significant decrease in soluble chitinase activity following inoculation in all clones except SNK13, SNK64, ICS95 and SNK413, where it remained almost constant. Between 5-day-old wounded pods and 5-day-old inoculated pods, no statistical differences were observed in chitinase activity due to inoculation. The only exception was clones SNK10, SNK213 and SNK64 where it decreased. Among 5-day-old inoculated pods, the highest soluble chitinase activity was recorded in clones SNK13 (0.712 units 10-4 g-1 fresh weight) and ICS95 (0.753 units 10-4 g-1 fresh weight), which was 318 % higher than the lowest activity (0.170 units 10-4 g-1 fresh weight) recorded in the clone ICS84 (Table 1). Ionically-bound chitinase activity in freshly harvested pods was higher in clones SNK10, SNK13, ICS95 and ICS84 than in the other clones (Table 2). The activity ranged from 0.129 unit 10-4 g-1 fresh weight (SNK416) to 0.250 units 10-4 g-1 fresh weight (SNK13). Five days after pod harvest, this activity increased in the pods of SNK213 (33%); decreased in clones SNK10 (25%), SNK13 (36%), ICS95 (19%), ICS84 (35%) and SNK413 (30%); while it remained almost constant in the clones UPA134, SNK64 and SNK416. When 5-day-old intact pods were compared with 5-day-old wounded pods, the activity of ionically-bound chitinase decreased in clones SNK10 (24%), UPA134 (43%), ICS95 (42%) and SNK416 (28%), while remaining almost constant in the other clones. Significantly high decreases were observed due to inoculation (for both intact and wounded pods) in clones SNK10, UPA134, SNK64 and ICS84. In 5-day-old wounded pods and 5-day-old inoculated pods, no statistical differences were observed due to inoculation in the activity of ionically-bound chitinase, except for clones SNK64, where it decreased and ICS95 and SNK416 where it increased (Table 2). The chitinase activity in inoculated pods was highest in clones SNK213 (0.158 unit 10-4 g-1 fresh weight) and this was 97% higher than the lowest activity (0.080 unit 10-4 g-1 fresh weight) measured in clone SNK64 (Table 2). Polyphenoloxidase (PPO) activity. Soluble PPO activity was higher in the freshly harvested pods of clones ICS95, SNK416 and SNK413 than in the six other clones (Table 1). PPO activity ranged from 13.28 DAbsorbance 30 s-1 g-1 fresh weight (clone SNK213) to 17.12 DAbsorbance 30 s-1 g-1 fresh weight (clone SNK413). In 5-day-old intact pods, PPO activity increased due to aging in all the clones, except SNK13, SNK213 and SNK416, where the activity remained constant. For the 5-day-old intact 5-day-old wounded pods, soluble PPO activity decreased in the clones SNK10 (20%) and SNK64 (35%), but remained constant in the other clones. In the case of 5-day-old intact and 5-day-old inoculated pods, there were statistical activity differences in clone SNK413 (where it increased) and in clones SNK64, ICS95, SNK416 and ICS84 (where it decreased). Between 5-day-old wounded pods and 5-day-old inoculated pods, significant differences due to inoculation were observed in the soluble PPO activity in clones SNK64 and SNK416 (where it decreased) and in clone SNK413 (where it increased) (Table 1). In inoculated pods, the highest soluble PPO activity was recorded in clone SNK413 (25.38 DAbsorbance 30 s-1 g-1 fresh weight), which was 103% higher than the lowest activity (12.48 DAbsorbance 30 s-1 g-1 fresh weight) in clone SNK13 (Table 1). For the ionically-bound PPO in freshly harvested pods, the activity was highest in clones SNK13 and SNK213. This activity ranged from 5.04 DAbsorbance 30 s-1 g-1 fresh weight (ICS84) to 14.60 DAbsorbance 30 s-1 g-1 fresh weight (SNK213). In 5-day-old intact pods, PPO activity increased in the cortex of ICS95 (22%) and ICS84 (30%), but decreased in the cortex of UPA134 (78%) and SNK213 (64%). It, however, remained constant in the cortex of SNK10, SNK13, SNK64, SNK416 and SNK413. Wounded pods showed that wounding did not affect the activity of ionically-bound PPO (Table 2). Native PAGE analysis of soluble isopolyphenoloxidases revealed two reproducible bands (A2 and A3) in all clones considered (Fig. 1). Clones ICS95 and SNK416 displayed one specific isoform (A1), which could be correlated with the aging of these clones and clone UPA134 displayed two specific isoforms (A4 and A5). For the ionically-bound isopolypheno-loxidases, band A'3 appeared independently in all cocoa pods (Fig. 2). The isoform A'1 was specific to the less susceptible clones, ICS84 and SNK413. Isoform A'2 was detected in freshly harvested pods of the highly susceptible clone UPA134 and in the infected pods of the less susceptible clones, ICS84 and SNK413. Isoform A'4 was specific to the freshly harvested pods of clone UPA134. DISCUSSION This study has shown that constitutively, the level of b-1,3-glucanase, chitinase and PPO activities in the different cocoa clones studied had no clear correlation with their susceptibility to P. megakarya. This finding is similar to that reported by Rosmin et al. (2000) on the same enzymes in relation to the susceptibility of cocoa to vascular streak dieback disease (Onchobasidium theobromae) and that reported on cucumber in relation to its susceptibility to Colletotrichum lagenarium (Irving and Kuc, 1990; Dalisay and Kuc, 1995). When plants are infected by microorganisms or damaged by mechanical injury, major physiological changes are induced and plant defence enzymes are generally activated. A role in plant defence mechanism has been assigned to b-1,3-glucanase and chitinase in a large number of species where pathogen induces earlier and higher increase in these enzyme activities in incompatible host-parasite interactions than in compatible ones (Caruso et al., 1999; Cordero et al., 1994; Ignatus et al., 1994). Our data demonstrate that in inoculated pods, soluble and ionically-bound b-1,3-glucanase activities were statistically highest in the less susceptible clones ICS84 and SNK413. The high accumulation of these hydrolases in the cortex of the less susceptible clones may play both a direct protective role by degrading fungal cell wall component or an indirect role in the plant defence mechanism by releasing some elicitors from the decaying fungal cell wall that stimulate phytoalexin accumulation in the host plant (Umemoto et al., 1994; Okinaka et al., 1995). In fact, it is well known that b-1,3-glucanase, in combination with chitinase release oligosa-ccharide, signal molecules that can activate a variety of plant defence events (Ryan, 1988; Krishnaveni et al., 1999). In our experiments, pod inoculation was generally followed by a decrease in chitinase activity in both soluble and ionically-bound fractions. This pattern did not correlate with cocoa susceptibility to P. megakarya. The same observation was reported by Irving and Kuc (1990) and Dalisay and Kuc (1995), who did not find a significant linear correlation between disease occurrence and the presence of chitinases in cucumber plants challenged with C. lagenarium. The decrease in chitinase activity on one hand, and the increase in glucanase activity observed only in the less susceptible clones, on the other, may at least in part, explain why all cocoa clones are susceptible to P. megakarya, but differ in level of tolerance to the disease. In fact, this difference is evident in the field whereby a low rate of disease development in the less susceptible pods compared with that in the highly susceptible pods which, are completely destroyed before maturity. It is apparent in this study that the change in PPO activity after pod inoculation was genotype-dependent. Polyphenoloxidase activity in the soluble fractions could not be correlated with the susceptibility of the different clones to P. megakarya. On the other hand, PPO activity in the ionically-bound fractions was characterised by an increase in the less susceptible clones SNK64, ICS95, SNK416, ICS84 and SNK413. Besides, some alterations were found when ionically-bound isopoly-phenoloxidases were analysed electro-phoretically after pod inoculation. The isoform A'2 appeared specifically in inoculated pods of the less susceptible clones ICS84 and SNK413. Similar observations are reported for mango (Medicago sativa) (Belluci et al., 1999) and cucumber (Cucumis sativus L.) (Chen et al., 2000) species upon attack by insect and Pythium aphanidermatum,respectively. Polypheno-loxidase is known to inhibit polygalacturonases of the pathogen (Stotz et al., 2000) and is involved particularly in the production of bactericidal and fungicidal hydroxyphenolics, and quinones that may polymerise to seal the infected tissue (Lazarovits and Ward, 1982; Vaugh et al., 1988). CONCLUSION Plants develop a complex variety of events that involve synthesis and accumulation of new proteins that can have direct or indirect action during pathogenesis. The coordinated induction of several PR proteins that may act synergistically is part of the defence strategy that plants activate against the invading pathogen and may limit the colonization of the plant by inhibiting pathogen growth. Cocoa pod inoculation with P. megakarya was characterised in our experiments by an increase in b-1,3-glucanase activity in the less susceptible clones and the appearance in these clones of an ionically-bound isopoly-phenoloxidase A'2. These alterations could be correlated to disease tolerance through the reinforcement of the lignification of the cell wall. Further assay involving cell wall extracts challenged by fungi would give more insight into the molecular interactions between T. cacao and P. megakarya. REFERENCES
©2003, African Crop Science Society The following images related to this document are available:Photo images[cs03012f2.jpg] [cs03012f1.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}