 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
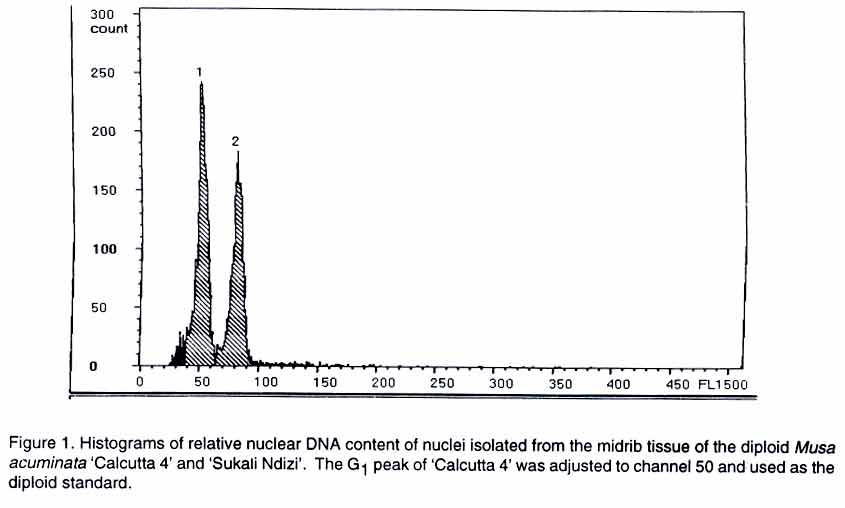
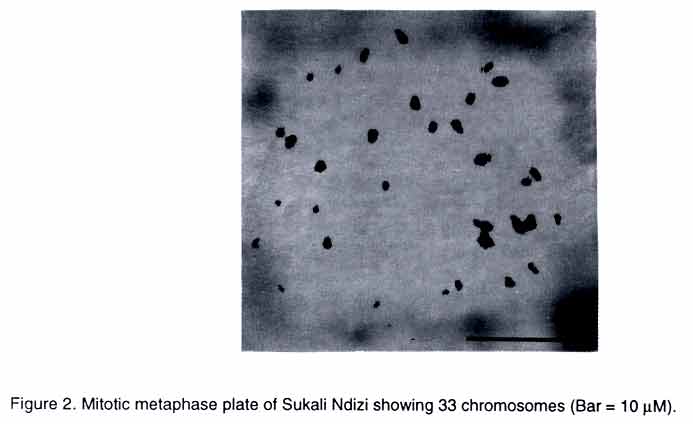
African Crop Science Journal, Vol. 11. No. 2, 2003, pp. 119-124 Short Communication ESTABLISHING THE GENOME OF 'SUKALI NDIZI' M. Pillay, J. Hartman*, C. Dimkpa and D. Makumbi International Institute of Tropical Agriculture, Eastern and Southern Africa Regional Center, P. O. Box 7878 Kampala, Uganda (Received 22 February, 2002; accepted 8 April, 2003) Code Number: cs03014 ABSTRACT Cultivated bananas originated from interspecific hybridisation of two wild diploid (2n = 2x = 22) species, Musa acuminata and M. balbisiana that were the donors of the A and B genomes, respectively. Most cultivated bananas are triploids with 2n = 3x = 33 chromosomes. They vary in genome composition with the most common types being AAA (dessert), AAB (plantain) and ABB (cooking banana). Banana improvement programs make use of interspecific hybridisation for gene introgression. Consequently knowledge of the exact ploidy level and genome composition of a plant is important for breeding purposes. The ploidy level of a plant is generally determined in two ways: (1) by a physical count of its chromosomes, and (2) by flow cytometry. In this study, the ploidy level of the dessert banana cultivar, 'Sukali Ndizi', was determined by conventional chromosome analysis and flow cytometry. Our results showed that 'Sukali Ndizi' is a triploid. In the past, 'Sukali Ndizi' was considered to be a diploid with an AB genome composition. We also determined the genome composition of the cultivar using a set of RAPD (random amplified polymorphic DNA) markers and found that it has an AAB genome composition. Key Words: Banana, interspecific hybridisation, ploidy level, RAPD RÉSUMÉ Les bananes cultivées obtenues par hybridation interspécifique de deux diploides sauvages (2n=2x=22), Musa acuminata and M. balbisiana étaient les donneurs des génomes A et B, respectivement. La plupart des bananes cultivées sont diploïdes avec 2n=3x=33 chromosomes. Elles varient en composition des genomes avec le type plus commun étant AAA (dessert), AAB (banane à cuirre). Le programme de bananes améliorées utilise l'hybridation interspécifique des génes introgression. Par conséquent, la connaissance du niveau exacte du poide et la composition des génomes de plantes est importante pour leur croisement. Le niveau de ploïdes dans une plante est généralement determiné dans de deux façons : par le decompte physique de leurs chromosomes, et par cytometrie. Dans cette étude, le niveau de ploïdes de la variété dessert de la banane, « Sukali Ndizi » était déterminé par l'analyse conventionelle des chromosomes et la cytométrie. Nos résultats ont montré que « Sukali Ndizi » est triploïde. Dans le passé, « Sukali Ndizi » était considéré comme un dipoïde avec un génome AB. Nous avons déterminé la composition génomique de la variété utilisant le marqeurs RAPD et avons trouvé que la variété avait une composition génomique AAB. Mots Clés: Banane, hybridation interspécifique, niveau du ploïde, RAPD INTRODUCTION Banana is a highly valued food crop in the highlands of East Africa. The region records one of the highest consumption figures for banana in the world at over 300 kg per capita /year (Karamura, 1998). There is a wide diversity of highland bananas in Uganda with over 80-recorded cultivars (Karamura, 1998). Until recently the taxanomy of bananas has been a big challenge to scientists in the region. However, it is now clear that the basic chromosome number in most cultivated and wild bananas is x = 11 (Stover and Simmonds, 1987). Cultivated bananas originated from interspecific hybridization of two wild diploid (2n = 2x = 22) species, Musa acuminata Colla and M. balbisiana Colla that were donors of the A and B genomes, respectively (Stover and Simmonds, 1987). Three main ploidy levels exist in Musa: diploid (2n = 2x = 22), triploid (2n = 3x = 33) and tetraploid (2n = 4x = 44). The highland bananas are triploid with 33 chromosomes and an AAA genome composition. Besides the endemic cultivars, there are a number of exotic or introduced cultivars in Uganda. One example is 'Sukali Ndizi'. It is characterized by having a compact bunch, short fingers and very sweet apple flavored fruit when ripe. It is used mainly as a dessert banana but also in juice preparations. 'Sukali Ndizi' is a widely distributed cultivar and is gaining importance in both local and export markets (Tushemereirwe et al., 2001). The plant has been considered to be a diploid with an AB genome composition (Karamura and Karamura, 1994). Like the other bananas in East Africa, 'Sukali Ndizi' is affected by a number of diseases and pests primarily black Sigatoka caused by Mycosphaerella fijiensis, Fusarium wilt, nematodes, weevils and viruses. Cross breeding of bananas is essential to produce improved hybrids that would be resistant to the various pests and diseases. Musa is a multi-ploidy crop and ploidy manipulation is critical to breeding. Crossing a triploid banana with diploid accessions generates diploid, triploid, tetraploid, aneuploid and hyperploid progeny (Vuylsteke et al., 1993). For selection of a hybrid of a desired ploidy level, there is need for precise determination of the ploidy of its parents. In addition, the genome composition of the hybrid will be determined by the genome contributions of its parents. Consequently knowledge of the exact genomes of a plant provides useful information to the breeder. DNA flow cytometry measures the fluorescence of a large number of stained nuclei within seconds and provides an estimate of nuclear DNA content within somatic plant tissues (Galbraith et al., 1983; Arumuganathan and Earle, 1991). The chromosome number of a plant is significantly correlated with nuclear DNA content (Verma and Rees, 1974; Black and Beckmann, 1983). Therefore DNA flow cytometry provides a much easier way of determining the ploidy level of a plant. MATERIALS AND METHODS Plant Material. Sukali Ndizi' clones collected from 16 districts in Uganda were used in this study. The 16 districts from which the clones were collected include: Lira, Soroti, Apac, Pallisa, Bushenyi, Kawanda, Mbale, Jinja, Kamuli, Kasese, Rukungiri, Mpigi, Rakai, Kibale, Mityana, Mbarara. The accessions are now located in the germplasm collection of the Kawanda Agricultural Research Institute, NARO, Uganda. The accessions were subjected to: (i) flow cytometry and conventional chromosome counting to establish the ploidy of 'Sukali Ndizi' and, (ii) genome specific RAPD markers to identify its genome composition. We have previously identified RAPD markers that are specific to the A and B genomes in Musa (Pillay et al., 2000). Details of the procedures followed in this study are as outlined in the sections that follow. Flow Cytometry. The cell nuclei were isolated by chopping a small piece of the midrib in a petri-dish with a sharp razor blade in 0.5ml of Otto buffer I (Otto, 1990). After adding an additional 0.5 ml Otto I buffer, the suspension was pipetted a few times to assist in releasing the nuclei. The nuclei were separated from the suspension by filtration through a 40 µM mesh nylon filter and stained by adding 1ml of Otto buffer II (Otto, 1990) containing DAPI (4',6-diamidino-2-phenylindole). The fluorescence of the nuclei was measured in a Partec (Partec, Munster, Germany) ploidy analyzer immediately after addition of Otto II buffer. The ploidy analyzer was calibrated so that the G1 peak of stained nuclei from M. acuminata cv 'Calcutta 4', a diploid, was set at channel 50. At least three different samples were measured for each plant and about 5000-10,000 nuclei were analyzed for each sample. Chromosome counts and genome composition. Chromosome counts from root tips of plants obtained from six of the sixteen locations were carried out as described by Pillay et al., (2001) and Pillay and Adeleke (2001). Briefly, fresh root tips were pretreated for 2 h in 0.036% 8-hydroxyquinoline and then fixed in 3:1 ethanol-acetic acid. The meristematic zones were digested at 37° C in an enzyme mixture consisting of 5% cellulase (Sigma Chemicals), 1% pectinase and 1% pectolyase Y23 (Karlan Research, Santa Rosa, Calif) made in a citrate buffer, pH 4.5. The enzyme solution was removed and the meristems were washed with water several times. A single meristem was placed on a glass slide, the excess water removed with a paper towel and 1 or 2 drops of freshly prepared 3:1 ethanol-acetic acid placed over it. The meristem was macerated and the cells smeared over the slide with a fine forceps. The slide was observed in a phase contrast microscope. When the cells began to adhere to the slide, several drops of the 3:1 was placed over one end of the slide and allowed to flow over the cells. The slide was air-dried and stained with Leishman's stain as described by Singh (1993). The RAPD markers that are specific for the A and B genomes of Musa (Pillay et al., 2000) were used to determine the genome composition of 'Sukali Ndizi'. The reaction mixtures for the RAPD analysis consisted of 0.2 µg DNA, 2.0 mM MgCl2 0.2 mM each dNTP, 1.25 U Taq Polymerase (Advanced Biotechnologies, Surrey, UK) and 1.2 µM of primer in a reaction buffer containing 75mM Tris- HCl pH 9.0, and 20 mM (NH4)2SO4 in a total volume of 25 µL. Three 10-mer primers A17, A18 and D10 (Operon Technologies, Alameda, Calif.) were used in separate reactions. Amplifications were done in a Perkin Elmer Cetus 9600 Thermocycler programmed as follows: an initial 3 min denaturation at 94°C followed by 35 cycles of 50s at 94°C, 50s at 40°C, and 1.5 min at 72°C, with a final extension of 7 min at 72°C. Approximately 15 µL of the amplification products were run on 1.2% agarose gels in 1X TBE buffer. RESULTS AND DISCUSSION Flow cytometry and chromosome counts. The nuclei isolated from the sixteen samples produced histograms at channel 75 (Fig. 1) corresponding to the G1 phase of the cell cycle. In higher plants the cell cycle is divided into three phases viz. G1, S and G2 (Howard and Pelc, 1953). The G1 phase of a diploid cell represents the normal or 2C nuclear DNA content (i.e., it possesses two copies of each gene). The DNA content of nuclei in certain phases of the cell cycle is, therefore, related to the ploidy level. The flow cytometer is used to measure the size of the nuclei and produce histograms that describe the DNA content of the cells. All the cells belonging to one peak have the same quantity of DNA and would represent a ploidy level. For example, all the nuclei of M. acuminata 'Calcutta 4' are identical in size and produced a histogram at channel 50 (Fig. 1). Therefore under standardized conditions, an unknown sample producing a peak at channel 100 would represent a tetraploid while that with a peak at channel 75 would represent a triploid. Flow cytometry showed that all the 'Sukali Ndizi' samples used in this study produced histograms with major peaks at channel 75 (Fig. 1) suggesting that it is triploid. Triploidy was confirmed by counting root tip metaphase chromosomes of plants from six of the sixteen locations. 'Sukali Ndizi' has green-a yellow, slender stem, with small fruits and stiff upright leaves and fits the morphological description of a diploid. This, however, is not the case and can be misleading. Chromosomes counts from root tips of 'Sukali Ndizi' from 6 of the 16 locations showed 33 chromosomes (Fig. 2) for each sample examined. Genome composition. Figure 3A shows that the 'Sukali Ndizi' clones shared the 600bp fragment that is present in M. acuminata (AA) and absent in M. balbisiana (BB). The 600 bp fragment is diagnostic for the A genome. Similarly, Figure 3B showed that the clones shared the 300 bp fragment with M. balbisiana, a fragment that is diagnostic for one B genome. Based on these results, it is probable that 'Sukali Ndizi' is a triploid with only one B genome and also has a band diagnostic for the A genomes, we assume that it has two A genomes and one B genome. Generally, ploidy and genome composition in Musa is assessed by scoring a number of morphological characteristics in mature plants. It is known that the environment influences morphological characters in Musa and this method can lead to inconsistent and unreliable results. For example, "Klue Tiparot" that was classified as a tetraploid (ABBB) on the basis of morphology is now confirmed to be a triploid with 2n = 3x = 33 chromosomes both by classical microscopy and flow cytometry (Jenny et al., 1997). Similarly, Horry et al. (1998) found that 'Pisang Jambe' and 'Kluai Ngoen' that were classified as a tetraploid and triploid, respectively, from their morphological behavior were actually a triploid and tetraploid, respectively. Careful screening of Musa genotypes with flow cytometric and molecular techniques will help to resolve similar existing problems. In this study, 'Sukali Ndizi' had the diagnostic band for the A genome (Fig. 3 A) and also had the diagnostic band for one B genome (Fig. 3B). Since flow cytometry showed that it is triploid, and DNA markers showed the presence of one B genome, we assume that the other two genomes are AA. CONCLUSION Two independent methods were used to determine the ploidy of 'Sukali Ndizi' in this study. Both conventional chromosome counting and flow cytometry showed that the plant is a triploid. This study also showed that 'Sukali Ndizi' has an AAB genome composition. Our findings are similar to those of other studies in Musa with regard to the determination of ploidy and genome composition in banana on the basis of morphology alone for which un reliable and in consistent findings have been reported (Jenny et al., 1997). ACKNOWLEDGEMENTS We are grateful to Dr. W.K. Tushemereirwe for granting us permission to use the 'Sukali Ndizi' plant collection at the Kawanda Agricultural Research Center. Funding for this research was provided, in part, by a grant from the Director General for International Cooperation (DGIC), Belgium. REFERENCES
©2003, African Crop Science Society The following images related to this document are available:Photo images[cs03014f1.jpg] [cs03014f2.jpg] [cs03014f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}