 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
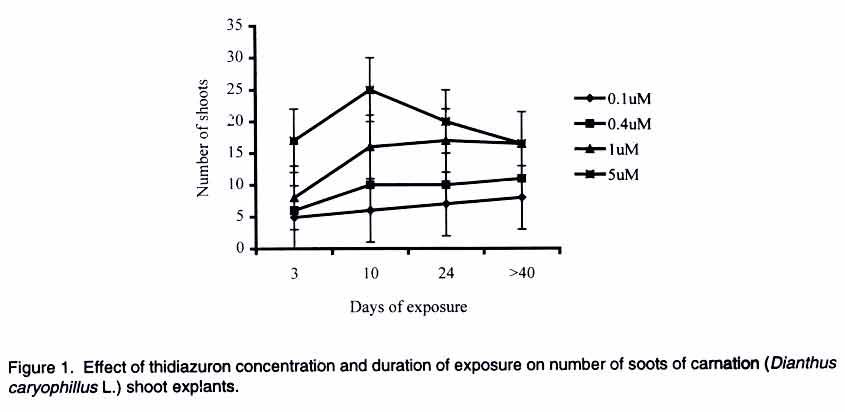
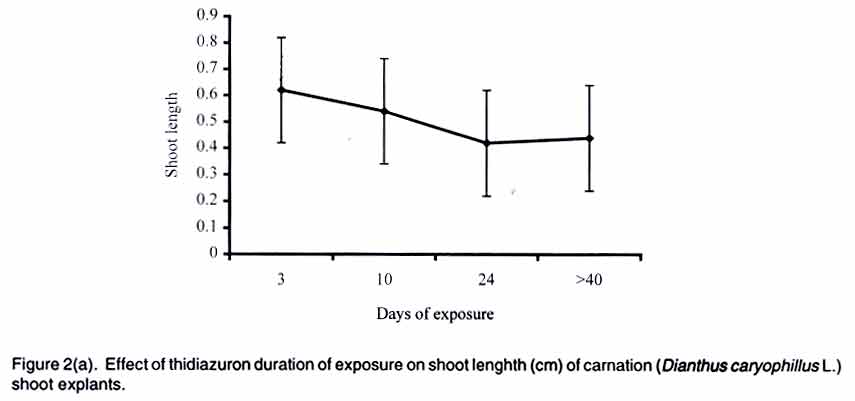
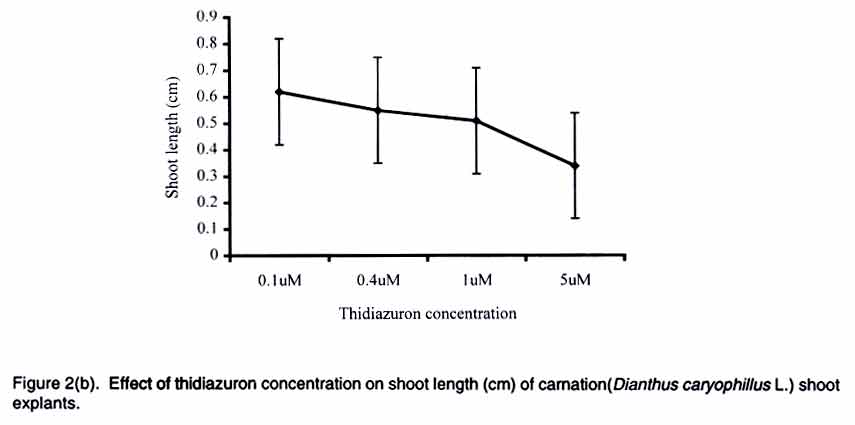
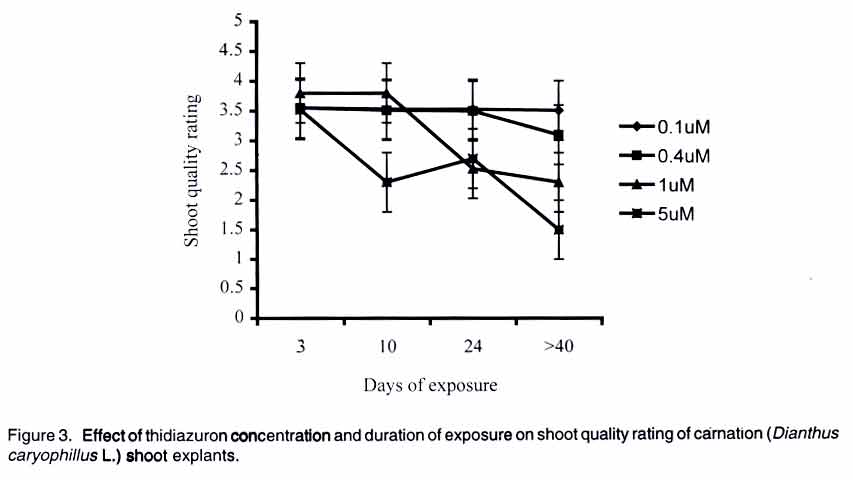
African Crop Science Journal, Vol. 11. No. 2, 2003, pp. 125-132 Short Communication EFFICACY OF THIDIAZURON IN In vitro PROPAGATION OF CARNATION SHOOT TIPS: INFLUENCE OF DOSE AND DURATION OF EXPOSURE R. Onamu, S.D. Obukosia, N. Musembi and M.J. Hutchinson Department of Crop Science, University of Nairobi, P. O. Box 30197, Nairobi, Kenya (Received 4 March, 2003; accepted 19 May, 2003) Code Number: cs03015 ABSTRACT One of the main challenges facing production of good quality cut flowers is the lack of clean planting material especially among resource poor farmers who resort to multiplying their own propagules with detrimental results. Carnation (Dianthus caryophillus L. cv. Yair) is an important export crop among farmers in Kenya. Carnation production, however, is constrained by lack of clean planting materials. The main objective of this study was to evaluate the potency of thidiazuron (TDZ), a phenyl urea, in evoking morphogenic response invitro compared to combined auxin (NAA) and cytokinin (kinetin) from shoot tip explants of carnations as one possibility of producing en mass clean planting materials. In addition, the study was instituted to determine the optimum concentration and duration of exposure of TDZ in the number, length and general quality of shoots formed. Results from the study indicate that TDZ, at low concentrations of 0.1-1 µM, was just as effective as 0.2 mg-1 L NAA + 0.2 m-1 L kinetin in improving the number, length and quality of shoots regenerated from the explants cultured in vitro. Although multiple shoots were formed from explants cultured on media supplemented with high doses of TDZ (5 µM), they were dwarfed and of a poorer quality. Prolonged exposure of carnation shoot tip explants to high concentrations of TDZ was detrimental to plant regeneration. Sufficient morphogenic response could be achieved by exposing the explants to 1-5 µM TDZ for short durations of 3-10 days with subsequent transfer to MS basal medium devoid of any plant growth regulators. Key Words: Cut flowers, Dianthus caryophillus, explants, Kenya, planting materials, plant regeneration RÉSUMÉ Un des principaux defis à la production d'une bonne qualité de coupe des fleurs est le manque d''n matériel propre adapté spécialement parmi les fermiers pauvres qui recourent à la multiplication de leurs propres propagules avec les détriments y résultants. L'oeillets (Dianthus caryophillus L. cv. Yair) est une importante plante d'export parmi les fermiers au Kenya. La production de l'oeillet cependant est contrainte par le manque des matériels adaptés de plantation. L'objectif principal de cette étude était d'évaluer de phenul, en evoquant une réponse morphogénique in vitro comparée au combiné d'auxin (NAA) et de cytokinin (Kinctin) à partir du bout du rejet des explants des oeillets comme une possibilité de produire en masse les matériels de plantation propres. En plus, l'étude était instituée pour déterminer la concentration optimale et la durée d'exposition du TDZ en nombre, la longueur et qualité générale des rejets formés. Les résultats de l'étude indiquent que les TDZ, aux faibles concentrations de 0,1-1 µM, était effective comme à 0,2 mg-1 L de Kinetin en améliorant le nombre, longueur et qualité des rejets r égénerés à partir des explants cultivés sur une couverture augmentée avec des doses élévées de TDZ (5 µM), ils étaient écrasés et d'une pauvre qualité. L'exposition prolongée du rejet d'oeillet à des fortes concentrations de TDZ était préjudiciable à la régéneration des plantes. La réponse morphogénique suffisante pourrait être atteinte par l'exposition des explants à 1- 5 µM TDZ pour les courtes durées de 3 - 10 jours avec transfert subséquent au basal moyen MS dénué de tout regulateur de croissance de plante. Mots Clés: Coupe des fleurs, Dianthus caryophillus, explants, Kenya, matériels de plantation, régénération de plantes INTRODUCTION Carnation (Dianthus caryophillus L.) is the second most important floricultural crop in Kenya, after roses (HCDA, 1997). Carnation has traditionally been propagated vegetatively by most Kenyan farmers, using cuttings from previous season's crop (Yako, 1999). This traditional method is both slow and ineffective, especially in the control of fungal and viral diseases in subsequent crops (Sparnaaij et al., 1991) leading to low yields and poor quality cut flower production (Wabule et al., 1991). Thus, the need to explore other more effective methods of propagation. Tissue culture offers an opportunity for farmers to access clean planting material and provides a good start in the production of good quality carnation cut flowers that can adequately compete in the international markets. Starting production of a crop using low quality planting materials negates benefits from other subsequent management practices. Most previous protocols that have been developed for in-vitro propagation of carnation from different explants, involve the use of combined auxin and cytikinin to induce morphogenesis (Frey and Janick, 1991; Frey et al., 1992; Nakano et al., 1994; Watada et al., 1996; Yanrcheve et al., 1998). Morphogenesis has been described as a process that occurs in three distinct stages of development, namely (a) acquisition of competence, (b) induction or determination, and (c) differentiation (Christianson and Warnick, 1988). The induction process on competent tissues leads to the formation of morphogenic cells, which become determined on the morphogenic pathway. The above separation is ultimately crucial in the scaling-up and mechanization of micro-propagation systems. Many propagation systems have not differentiated these stages of development. For example, maintaining hypocotyls explants on 0.4 µM thidiazuron (TDZ), for the entire period of somatic embryigenesis is limited in indicating the optimum level of and duration of exposure to TDZ required for adequate stimulus or induction of the process (Visser et al., 1992). Further work by Hutchison et al. (1996a) indicated that the embryogenic process could be compartmentalized into a 3-day induction phase when explants are maintained on an induction medium containing 10 or 20 µM TDZ and an expression of growth/expression phase when explants are placed on a basal medium devoid of any plant growth regulators. Continuos exposure of explants to high concentration of TDZ is reported to be inhibitory/toxic, while low concentrations may not provide adequate stimulus (Huetterman and Preece, 1993; Lu, 1993; Hutchison et al., 1996a; Murthy et al., 1995). No research findings have been reported on optimum concentrations and durations of exposure of TDZ in carnation explants cultured in vitro. Thidiazuron, (N'-phenyl-N'-1,2,3-thidiazol-5-ylurea, TDZ), is a phenyl urea that was originally developed as a cotton defoliant but has gained importance as a potent plant growth regulator for in vitro propagation systems of various crops (Fiola et al., 1990; Visser et al., 1992; Murthy et al., 1995; Hutchison et al., 1996a; b). Despite its high efficacy in inducing morphogenic responses in vitro, there are limited reports where TDZ has been used to regenerate carnation. Better/more effective regeneration was obtained from carnation cv. White Sim shoot explants cultured on a membrane raft floating on liquid medium compared to those on agar-gelled or shaken liquid medium (Watada et al., 1996). Thidiazuron was reported to cause greater shoot regeneration from petal explants of carnation cv. Scania than kinetic, BAP or 2iP (Nakano et al., 1994). Interestingly, maximum shoot proliferation was achieved when petal explants of carnation cv. Scania, Improved White Sim and Sandra were cultured on MS medua supplemented with 0.05 µM TDZ and 0.5 µM NAA (Frey and Janick, 1991). Based on review of available literature, no research findings have been reported on the role of TDZ on morphogenesis of caranation cv. Yair, a popular cultivar among Kenyan flower farmers (Wabule et al., 1991). This study therefore was instituted to evaluate the potency of TDZ in evoking morphogenes in response of carnation explants in vitro. MATERIALS AND METHODS Stock plant care. Carnation (Dianthus caryophillus L.) source plants were obtained from a farmer's field in Kiambu district of Kenya, which is situated at 2100 m above sea level and around 10o South of the Equator. The carnation plants were planted and cultured under optimal cultural conditions for spray carnations. Explant preparation and surface sterilisation. Carnation shoots from 11/2 year old plants and having 12 nodes were cleaned with detergent and rinsed in running water for 15 min. Excised tips (1-2 cm long) were placed in sterile water in a beaker. The tips were surface sterilized by immersing in 70% ethanol for 5 min and then immersing in 1.5 % sodium hypochlorite solution containing between 20 (2 drops/100 ml solution) for 5 mins and then rinsed three times with sterile distilled water. Shoot tips consisting of an apical and one or two leaf primodia and measuring between 0.5 - 1.0 mm long were excised under a dissecting microscope and used as explants. Culture of shoot tip explants. One excised shoot tip was cultured aseptically in a universal bottle containing 10 mL of media. The culture medium consisted of MS (Murashige and Skoog, 1962) salts, B5 (Gamborg et al., 1968) vitamins, 30 g/L sucrose and was supplemented with various plant growth regulators in various concentrations and/or combinations of (a) TDZ (0.1, 0.4, 1 and 5 µM), (b) TDZ = NAA: (0.1 µM TDZ + 0/01mg/L NAA, 1µM TDZ + 0.01 mg/L NAA, 1 µM TDZ + 0.1 mg/L NAA), (c) 0.2 mg/L kinetin + 0.2 mg/L NAA (The concentrations used in the present study was determined using a cytokinin/auxin grid (Kyte, 1992) and based on previous studies/ procedures (Zimmerman et al., 1986; Mujib and Pal, 1994) and our preliminary studies on the crop, and (d) Basal medium devoid of any plant growth regulators acted as a control in all experiments. The explants cultured in media supplemented with TDZ were maintained on the media for 10 days and then transferred to MSO. All other explants were maintained on the medium for 4 weeks and sub-cultured to fresh similar medium after every 4 weeks. For the second experiment on the effect of dose and duration of TDZ on morphonogenesis from carnation shoot tips, explants were cultured on various concentrations of TDZ (0.1, 0.4. 1, 5 µM) for different periods (3, 10. 24, 30 days) and subsequently transferred to basal medium devoid of any plant growth regulators. The pH of the media were adjusted to 5.7 + 0.1 before autoclaving at 121oC for 20 min. The cultures were placed on growth shelves set at 25 + 2oC and illuminated (16 h photoperiod 70 - 78 µmo/m2/s) by cools white fluorescent tubes. Shoot length, numbers of leaves/stem and the shoot quality rating were assessed every week for a period of 3 months (12 weeks). Shoot quality rating involved a visual rating of 1-4; where 1 = poor growth, low chlorophyll content and browning of tissues; 2 = pale green shoots showing symptoms of severe chlorosis and fair to poor growth; 3 = light green shoots showing symptoms of slight chlorosis and good growth; and 4 = green healthy shoots with vigorous growth. Experimental design and statistical analysis. All experiments were laid out in a Completely Randomised Design and each treatment was replicated 3 times. The experiments were repeated twice. Data were analyzed using the analysis of variance (GENSTAT) statistical package (Lane and Payne, 1996) and the means were compared using Tukey procedure at 5% level of significance. RESULTS Addition of various plant growth regulators to the culture medium of carnation (Dianthus caryophillus L. cv. Yair) shoot tip explants, significantly influenced the number, length and general quality of shoots after 4 weeks in culture. Inclusion of TDZ or 0.2 mg/L NAA + 0.2 g/L kinetin significantly increased the number of shoot/explant (Table 1). The regeneration of shoots increased with increasing levels of TDZ in the culture medium (Fig. 1), the greatest number of shoots (32) being formed on 5 µM TDZ compared to 1 shoot on basal medium (Table 1). Although the shoot tips cultured on media supplemented with low concentrations of TDZ (0.1-0.4 µM) and 0.2 mg/L NAA = 0.2 g/L kinetin were fewer in number, the shoots were longer and of better quality than those formed on media supplemented with 5 µM or basal medium (Table 1). In addition to multiple shoot formation by explants cultured on 5 µM TDZ, the explants formed friable to compact, pink to green callus (Data not shown). The dose and duration of TDZ in culture medium had a significant effect on the number of shoots, shoot length and shoot quality. The number of shoots per explant increased with prolonged duration at low concentrations of TDZ (0.1 µM). At moderate concentrations (0.4-1 µM), the number of shoots increased with increasing concentration up to 24 days of exposure with longer periods (>40 days) either having no significant further increase or causing a slight reduction (Fig. 1). At 5 µM concentration, TDZ showed the greatest number of shoots with increasing duration up to 10 days. Prolonged duration of exposure to TDZ, in general, significantly reduced the number of shoots formed per explant (Fig. 1). The shoots cultured on various TDZ concentrations were significantly longer when exposed for short durations (Figure 2a) and low concentrations (0.1-1 µM, Figure 2b). Prolonged exposure (>40 days) of explants to high doses (5 µM) of TDZ significantly reduced shoot length of the regenerants (Fig. 2a-b). Better quality shoots were formed when explants were exposed to various concentrations of TDZ for a short duration of 3 days (Fig. 3). Increasing exposure beyond the 3 days significantly reduced the quality of shoots formed from shoot tip explants (Fig. 3). Exposing explants to 1 or 5 µM TDZ for longer periods (>10 and 24 days, respectively) significantly reduced the quality of shoots (Fig. 3). Low doses of TDZ promoted formation of fairly good quality shoots (>even with prolonged exposure >490 days) (Fig. 3). DISCUSSION AND CONCLUSION Thidiazuron is a phenyl urea that has recently gained importance as being more or just as potent as combined auxin and cytokinin in evoking morphogenic responses in vitro (Mok et al., 1982). Traditional methods of regenerating plants in vitro involve the use of auxins and cytokinins either alone or in combination (Skoog and Millet, 1957). Research on various plant regeneration systems indicate that morphogenesis is regulated by an intricate balance of endogenous auxin and cytikinin (Skoog and Miller, 1957, Hutchinson et al., 1996a). The results from the present study show that morphogenesis from carnation (Dianthus caryophillus L. cv. Yair) shoot tip explants occurred when explants were cultured in media supplemented with either auxin + cytokinin or TDZ. Thiadiazuron at low concentrations was as potent as tested combined NAA and kinetin in terms of number, length and general quality of shoots regenerated. Higher concentrations of TDZ (5µM) increased multiple shooting but had a dwarfing effect on the shoots, an observation consistent with work done previously on maize, oats, radish (Devlin et al., 1989), sour orange (Pountney and Swietlik, 1988), and other plant systems (Huetterman and Preece, 1993; Lu, 1993). Inhibitory effect of TDZ could be attributed to competition of nutrients among multiple shoots formed (Salisbury-Ross, 1991) or to cytokinin-like effects (Vanderhoef et al., 1973; Beyer et al., 1984). Cytokinins have been classified into adenine and phenylurea derivatives depending on their chemical structures (Mok et al., 1987; Takahashi et al., 1978) and thidiazuron belongs to the latter group. Thidiazuron, at high concentrations, has been shown to stimulate ethylene production (Suttle, 1985, Thomas and Katterman, 1986, Hutchison et al., 1996a,b). Regenerated shoots of carnation shoot tip cultures placed on 5 µM TDZ were more pale in colour than those cultured on low concentrations of TDZ or combined NAA and kinetin. Previously, yellowing of carnation petals was reported by Eisinger (1983). Our results however, are in contrast with reports indicating increased chlorophyll synthesis by phenyl ureas and other cytokin or cytokinin-like compounds (Kaul and Sabharwal, 1972; Kefford et al., 1973; Lew and Tsuji, 1982). Prolonged exposure of various plant explant to high concentrations of TDZ has been widely reported to be detrimental to morphogenic responses in vitro (Mok et al., 1982; Huetterman and Preece, 1986; Murthy et al., 1995). Morphogenesis has been shown to consist of 3 phases, the last two being induction and growth and development, respectively (Christianson and Warnick, 1988; Hutchison et al., 1996a,b). Results from the present study indicate that the length of induction period and its distinction from the development phase in morphogenesis of carnation shoot tip explants is dependent on the concentration of TDZ in the culture medium. Optimum response was obtained when explants were cultured for 3-10 days in 1 µM TDZ with subsequent transfer to basal medium devoid of any plant growth regulators, suggesting that the induction period is about 10 days followed by an expression in each development phase in the absence of TDZ. Prolonged exposure to TDZ was detrimental to shoot length and quality. In conclusion, results from the present study indicate that TDZ at low concentrations of not more than 1µM, is just as potent as combined auxin (NAA) and cytokinin (kinetin). In addition, shoot tip explants can be cultured for a brief period of 3-10 days in 1-5 µM and subsequently transferred to an MS basal medium devoid of any plant growth regulators for optimum morphogenic response in carnation (Dianthus caryophillus L) cv. Yair shoot tip explants. REFERENCES
©2003, African Crop Science Society The following images related to this document are available:Photo images[cs03015t1.jpg] [cs03015f3.jpg] [cs03015f2a.jpg] [cs03015f2b.jpg] [cs03015f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}