 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 11. No. 2, 2003, pp. 133-142 SHORT COMMUNICATION RESPONSE OF TOMATO GENOTYPES TO INDUCED SALT STRESS S.G. AGONG1,2, M. KINGETSU1 , Y. YOSHIDA1, S. YAZAWA3 and M. MASUDA1
1Department of Eco-physiology for Crop Production, Faculty of Agriculture,
Okayama University,
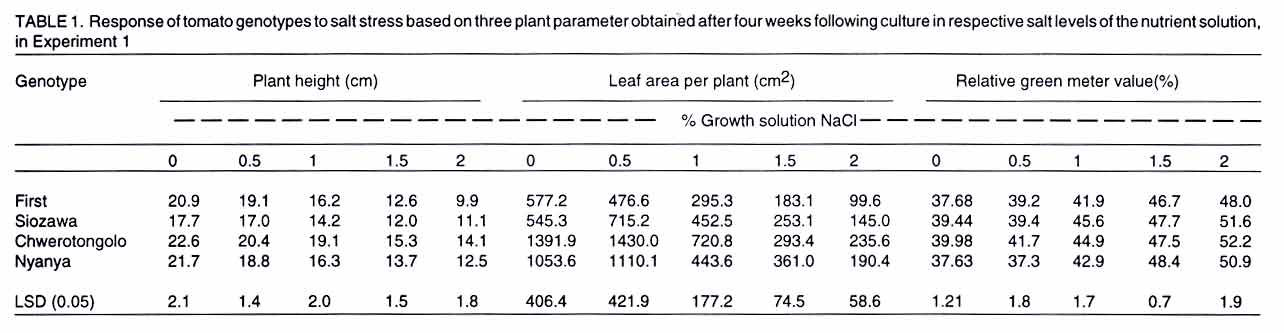
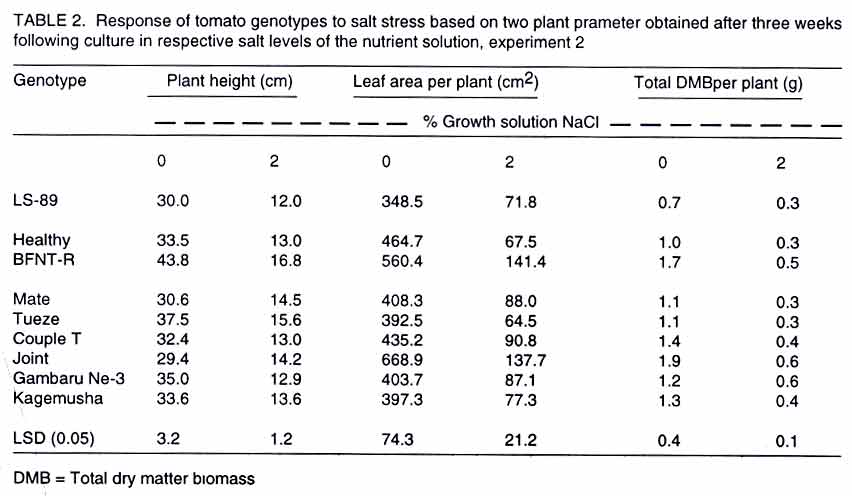
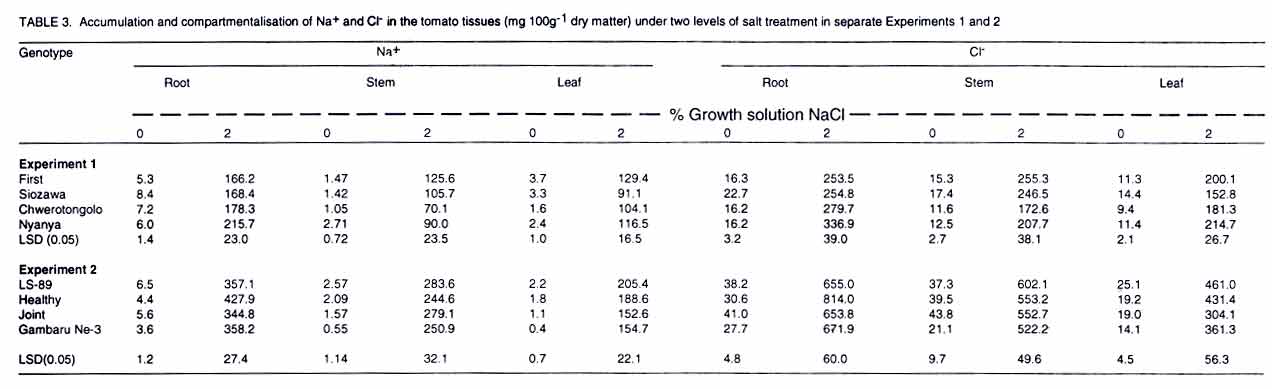
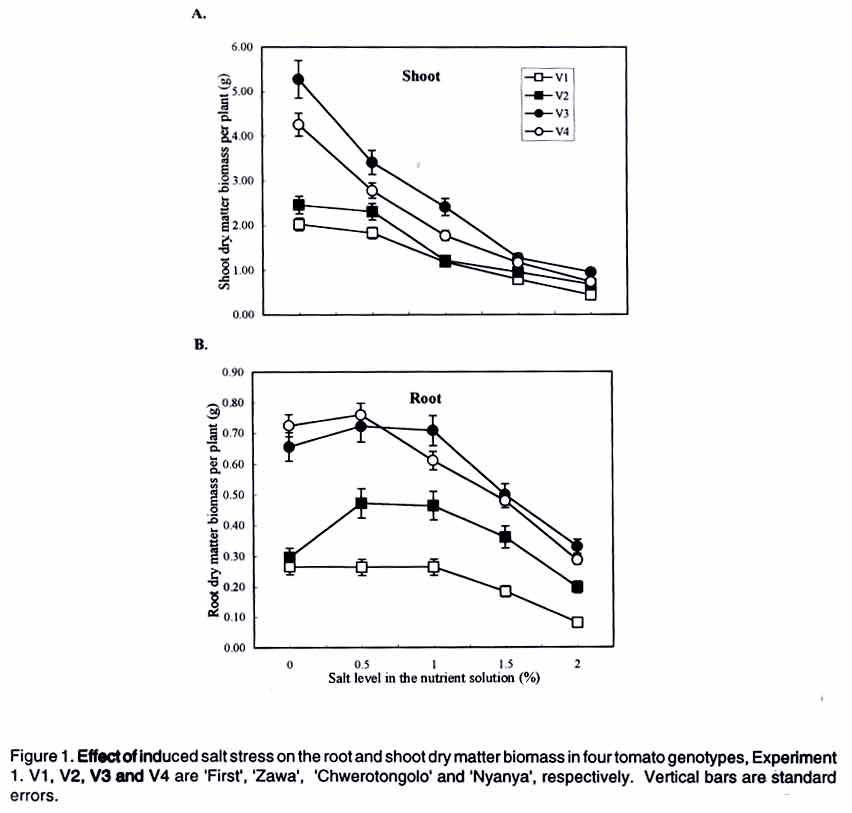
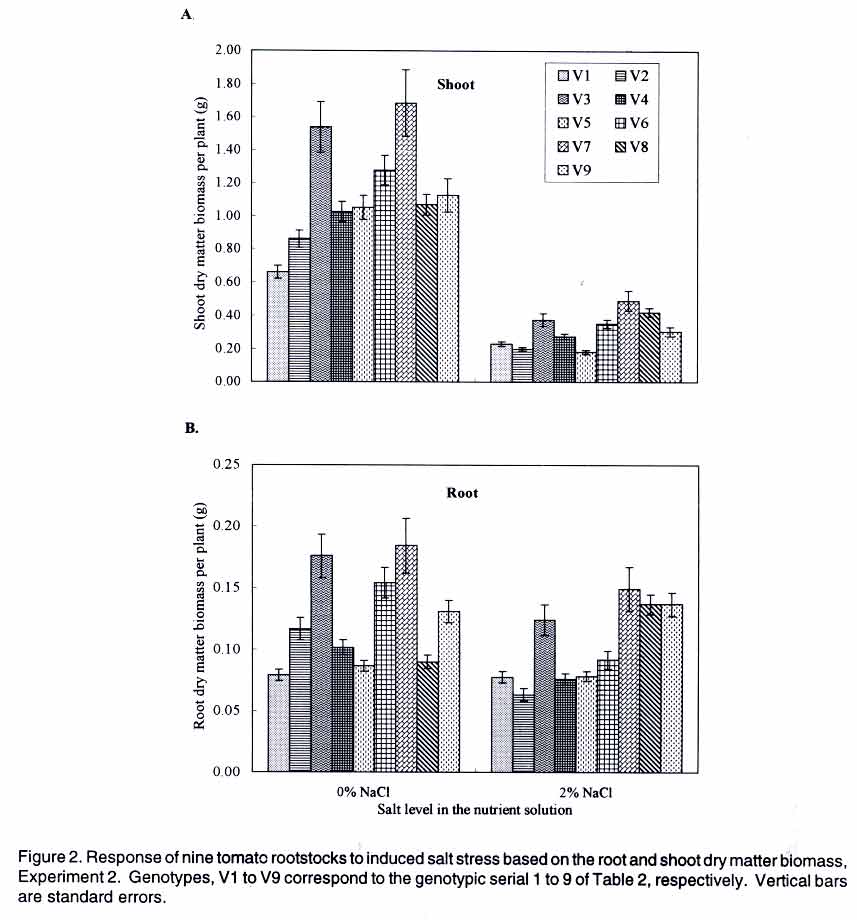
Tsushima-naka, Okayama 700-8530 Japan (Received 7 February, 2000; accepted 31 March, 2003) Code Number: cs03016 ABSTRACT Thirteen tomato (Lycopersicon esculentum L.) genotypes were subjected to salt treatment under hydroponics and their responses monitored in a set of two experiments with the objective of advancing them as potential salt tolerant tomato scion and/or rootstocks. Salt applications ranged from 0 to 2% NaCl, with the resultant EC values of 1.4 to 37 dS m-1, respectively. Genotypes were cultured in the experimental solutions for up to four weeks in the greenhouse. Significant genotypic and/or salt treatment effects were registered on plant height, leaf green meter value and area, dry matter yield, Na+ and Cl- accumulation in tomato tissues. Salt treatment at 2% NaCl stimulated chlorophyll production, but caused severe depression on dry matter yield and leaf area. Some tomato genotypes consistently showed superior biological activity at higher salinity and others exhibited greater shift in the shoot:root ratio (from 8:1 to 5:1 for 'First'), based on dry matter biomass production thus displaying relatively greater adaptation to salt stress. Two tomato genotypes ('Siozawa' and 'Gambaru Ne-3') displayed superior performance. Key Words: Genotypic variation, hydroponics, rootstock, salinity, shoot:root ratio RÉSUMÉ Treize genotypes de la tomate (Lycopersicon esculentum L.) étaient traités au sel dans des conditions hydroponiques. Leurs réponses étaient observées dans un ensemble de l'expérience avec comme objectfs d'améliorer les tomates comme scion partiellement tolérant au sel/ ou rootsocks. Les taux d'application rangés entre 0 et 2% de Nacl, résultant à des valeurs de EC rangeant entre 1,4 et 37 d S/m, respectivement. Le sgénotypes étaient cultivés dans de solutions experimentales jusqu'à quatre semaines dans le serre. Les effets génotypiques et/ou salin significatifs étaient enregistrés à la hauteur de la plante, la longueur et la surface de la feuille verte, le rendement en matière sèche, l'accumulation dans les cellules de la tomate du Na+ et cl-. Le taux de 2% de Nacl a stimulé la production du chlorophyle, mais a causé une chute dans le rendement en matière sèche et la surface verte de feuilles. Certaines tomates ont montré de manière consistente des activités biologiques pour des taux de sel élévés. En se basant sur la production de la matière sèche, d'autres ont exhibé de grandes variation dans le taux rejet-racines (de 8 :1 à 5 :1), par conséquent ont montré une adaptation relativement grande aux stress causés par le sel. Deux génotypes de la tomate ('Siozawa et Gambaru Ne-3) ont montré des performances élévées. Mots Clés: Variation génotypique, hydroponique, stock des racines, salinité, taux rejet-racine INTRODUCTION Most cultivated plants, tomato (Lycopersicon esculentum) inclusive, are sensitive to salinity stress (Mass, 1986). However, tomato production has been gradually extended into the more marginal lands, thus, exposing the crop to a greater risk of salt stress (Agong et al., 1997). On the other hand, tomato is a popular vegetable necessitating its improvement to fit in the salinity prone marginal areas in order to increase more production. For increased production of the tomato crop under saline environment, suitable cultivars are required to overcome the serious limitation posed by the vast salt affected semi- and arid-lands. The quest for better tomato yielding varieties for the marginal areas continues to receive global attention with limited breakthrough in producing salt tolerant tomato cultivars (Rush and Epstein, 1981; Saranga et al., 1991, 1992; Asins et al., 1993a,b; Alarcón et al., 1994; Adams and Ho, 1995; Bolarín et al. 1995; Cayula et al., 1996; Copeman et al., 1996; Pérez-Alfocea et. al., 1996; Agong et al., 1997; Foolad and Li, 1997; Foolad and Chen, 1998; Cuartero and Fernández-Muñoz, 1999). In their previous study, Agong et al. (1997) observed a few landraces to be fairly tolerant to NaCl treatments, unlike some market cultivars, on the basis of relative dry matter biomass. Among the landraces, 'Chwerotonglo' was consistently stable in terms of fruit dry matter production at moderate salinity, and had relatively higher fruit dry matter content. Nonetheless, due to relatively low fruit yields, this accession requires improvement before it can be advanced to the stakeholders for production. Improving tomato yield in a relatively salt tolerant cultivar will lead to degeneration of the salt tolerance ability in the progenies derived from the hybridization. Thus, tolerant low yielding tomato genotypes are unsuitable candidates for direct tomato yield improvement. In view of this dilemma, it follows that the priority of research with respect to salt tolerance in tomato should be re-examined with an ultimate goal of developing relatively suitable strong tomato salt tolerant rootstocks. This will not only facilitate exploitation of the more salt affected soils, but also strongly promote higher tomato productivity. Similarly, dependable high yielding tomato scions that easily form graft union with the salt tolerant rootstocks should be identified. The rootstock development is of practical inclination in solving the problem posed by the salinity with a clear vision of directly improving tomato productivity. Generally, it is well documented that true salt stress tolerance does not exist in the tomato cultigens (Foolad and Chen, 1999) except for the wild relatives (Maas, 1986). It is also believed that the crop and its relatively tolerant hybrids show more of adaptability than tolerance with substantial reduction in economic yields (Agong et al., 1997). Therefore, the main objective of this study was to evaluate the suitability of thirteen tomato genotypes for salt tolerance with an aim of advancing them as rootstocks for salt prone areas, under relatively moderate salinity. Verification of the possible practical utility of the landraces like 'Chwerotonglo' and 'Nyanya' in the improvement of tomato for salt tolerance and simultaneous comparison of these genotypes with two chosen Japanese cultivars ('Siozawa' and 'First') formed the core of the first experiment. In the previous study, Agong et al. (1997) concluded that improvement of the tomato crop should be possible with landraces playing the role of the base material. However, we believe that practical improvement strategy should revolve in the development of strong rootstocks for salinity tolerance. This strategy will be a fundamental remedial step in counteracting the salinity stress and enhancing economic yield of tomato under the harsh marginal water quality and quantity. Furthermore, the genetic control of the salt tolerance has proved to be extremely difficult to elucidate and remains elusive in the near future for practical utilization (Asins et al., 1993a). Subsequently, in our second experiment, we investigated the ability of nine tomato rootstocks with various kinds of soil-born disease resistance, to tolerate salinity with the objective of advancing them as potential materials for tomato production in salt affected lands. MATERIALS AND METHODS Experiment 1. Tomato genotypes 'First' (V1), 'Siozawa' (V2), 'Chwerotonglo' (V3) and 'Nyanya' (V4) were submitted for the salinity tolerance investigations. 'First' is a popular genotype widely used in Japan as a breeding line but its ability to tolerate salinity stress has not been established. On the other hand, accession 'Siozawa', a tomato genotype originally developed in Japan, is known for tolerating relatively high levels of salinity. The Kenyan landraces 'Chwerotonglo', and 'Nyanya' were similarly used in this study due to their dimorphic variation for salt stress tolerance (Agong et al., 1997). Salt susceptible and tolerant tomato cultigens of both the Japanese and Kenyan origin were utilised as the experimental plant materials. Tomato seeds were initially germinated in open flat plastic trays containing vermiculite, on the 30 August 1999 in the green house and raised for two weeks before being transferred to various salt stress treatments. The seedlings were watered with a half-strength Enshi nutrient solution (Hori, 1966). The solution comprised of: KNO3, Ca(NO3).4H2O, NH4H2PO4, MgSO4.7H2O in the proportion of 0.81, 0.94, 0.15 and 0.49 g L-1 as at full strength, respectively. The micronutrients were F-EDTA, H3BO3, MnCl2.4H2O, ZnSO4.7H2O, CuSO4.5H2O and NaMoO4.2H2O, with Fe, B, Mn, Zn, Cu and Mo amounting to 0.5, 0.5, 0.05, 0.02 and 0.01 ppm also as at full strength, respectively. Two week-old tomato seedlings were subjected to the salt stress in a hydroponic system for four additional weeks after which the plants were harvested. Salt treatments for the hydroponic culture were achieved via gradual addition of NaCl to the nutrient solution. The five levels of salt were: 0 (control without addition of salt), 0.5, 1, 1.5 and 2% NaCl. Except for the control, the rest of the treatments received salt on every other third day commencing two days after exposure only to the basal nutrient solution. Thus, a part from the necessary nutrients, the control solution was devoid of the NaCl. All the other treatments received appropriate amounts of salt in addition to the nutrients. The resultant electrical conductivities obtained following gradual salt addition of 0, 0.5, 1, 1.5 and 2% NaCl were 1.5, 12.8, 21.56, 28.9 and 38.5 dS m-1, respectively. A 20-litre volume of hydoponic bathing nutrient solution was maintained through addition of 1-2 liters of the nutrient solution every third day to make up for the lost water till the full salt gradients were attained after which the plants only received irrigation water to compensate for evapotranspiration. At harvesting, data were scored on plant height (cm), relative chlorophyll content in youngest fully expanded leaves; leaf area per plant; and dry matter biomass of root, stem and leaf. Also, from the dry matter biomass of the various tomato tissues (leaf, stem and root), the contents of Na+, and Cl- were determined as detailed below. Initially, the dried tissue was finely ground using a vibrating sample mill (Model T1-100), followed by extraction of the two minerals from a 100-mg sample of the tissue in 20 ml distilled water under boiling temperatures for 3 hours. The cooled individual samples were then filled to 50 ml with distilled water and shaken thoroughly before filtration. About 10 ml of the sample was filtered and stored under freezing temperatures for one week during which analysis was performed. Just before analysis, the samples were subjected to a re-filtration using a syringe fitted with a 0.45-µm pore diameter. The fine filtrate was analysed for Na+, Cl- and K+ contents using ion chromatographic procedure (Shimadzu CDD-6A, Conductivity Detector). Separation for both Na+ and K+ analyses was accomplished on Shimadzu cation column, with a 2.5 mM oxalic acid dihydrate as the eluant. That of Cl- proceeded on Shimadzu anion column with 2.5 mM phatalic acid as the mobile phase. The samples were run at a constant temperature of 40 C. Calibration solutions constituted from NaCl comprised of 5, 10 and 20 ppm and corrected for the individual minerals as appropriate. Potassium was calibrated using 10 ppm KCl solution. The experimental design used was a three replicate completely randomized block arrangement. In each treatment combination, four plants were studied. The data generated were subsequently subjected to analysis of variance (ANOVA) and mean separation was done using least significant difference (LSD0.05) procedure. Experiment 2. In this experiment, nine commonly utilised tomato rootstocks in Japan, with known pathological resistance, were investigated for ability to tolerate salt stress. Cultural procedures remained unchanged as in experiment 1, except that the seedlings of these rootstocks were planted two weeks later and harvested one week earlier. Also, only two levels of salt stress were used, namely: 0% (control) and 2% NaCl. The experimental design and replications were retained as in Experiment 1. Similarly, data collection and analysis proceeded as already outlined above. RESULTS There were significant (P<0.05) differences among the tomato genotypes as well as salt effects (Table 1). Consistently, salt treatments exerted depression on most plant parameters studied, irrespective of the genotypes evaluated. Plant height, green meter value and leaf area were significantly variable among the salt level across the tomato genotypes in Experiment 1 (Table 1) as well as the rootstocks in Experiment 2 (Table 2). Similarly, Na+ and Cl- accumulation in the plant tissues was significantly (P<0.05) influenced via salt stress across genotypes and salt levels (Table 3). The severe reduction in plant height demonstrated a consistent effect of salt stress in plant growth rate across the salt levels. In Experiment 1, Chwerotonglo was superior with respect to plant height and was visually relatively stable and healthier even under the highest salt treatment (2% NaCl). Similarly, 'BFNT-R' exhibited better visual performance as well as plant height among the nine tomato rootstocks (Table 2). Moderate salinity ( 0.5%) stimulated expansion of leaf surface in 'Siozawa' and maintained leaf surfaces comparable to the control in the case of 'Chwerotonglo' and 'Nyanya' (Table 1). Noticeably, steady reduction in leaf area occurred in 'First'. Salt treatments above 0.5% caused serious leaf surface reduction in all the genotypes, with the genotypes that relatively had lower leaf area as at control showing lesser fall in the final leaf area as at 2%. Overall, the Kenyan based tomato genotypes exhibited a reduction in leaf area. Nonetheless, also insignificant salt stress effects on the leaf area were recorded at lower salt level (0.5%) for 'Chwerotonglo', the 'Nyanya' and 'Siozawa', with the exception of First variety. The observation suggested a physiological advantage of the former three genotypes over the latter, at relatively low salinity. Further analysis of the leaf area data suggested a systematic osmotic adjustment occurring in the tomato genotypes, while under the exposure to the salt treatments via the tremendous reduction in the leaf area per plant (Table 1). Consequently, the plants with heavily reduced leaf area could only afford much less biomass production as was observed in the rootstock (Table 2). On the other hand, green meter values did not offer obvious differences among the tomato genotypes, but clearly demonstrated a steady adjustment in response to salt stress via elevated values with the increasing salinity across the four genotypes examined in Experiment 1 (Table 1). Analysis of root and shoot dry matter biomass in Experiment 1 suggested a high susceptibility of the four tomato genotypes to salinity under the hydroponic system (Fig. 1). Notably, salt treatment stimulated the production (or maintenance) of more roots at a moderate salinity of 0.5 to 1% NaCl, with 'Siozawa' showing the greatest response. Despite their high susceptibility to salinity, 'Chwerotonglo' and 'Nyanya' apparently exhibited very high genetic potential as directly discernible from the superiority expressed across the salt level compared to the other two genotypes. In Experiment 2, similarly, there was a considerable decrease in the root and shoot dry matter biomass across the genotypes at 2% NaCl treatment (Fig. 2). 'BFNT-R' and 'Joint' exihibited superior performance based on both root and shoot dry matter biomass per plant. On salt treatment, dry matter production was adversely reduced in all the nine rootstocks, except for 'Gambaru-Ne 3' and 'Kagemusha'. In both experiments, the shoot component was most severely affected by the elevated salinity treatment, with less perturbation on the root proportion. For instance, 'Gambaru Ne-3' significantly produced more root dry matter biomass at higher salinity treatment. Relative dry matter yields, derived from both experiments, revealed that 'Siozawa' and 'Gambaru Ne-3' were more tolerant to salt stress than the other genotypes in both experiments, respectively (Figs. 1 and 2). In the first experiment, 'Siozawa' was considered relatively tolerant to the salt stress, but, 'First', 'Chwerotonglo' and 'Nyanya' were not significantly different on the basis of relative dry matter biomass. Based on this study, the three genotypes are regarded as relatively weak to the induced NaCl stress. Observations made in Experiment 2 placed 'Gambaru-Ne 3' and 'Tueze' as relatively tolerant and weak tomato rootstocks, respectively (Figs. 1 and 2). The correlationship between root and shoot dry matter biomass investigated in the two experiments independently, revealed different biological activities in the tomato genotypes (Figs. 1 and 2). Overall, more root than shoot production occurred across the genotypes at the higher salt treatment, with each tomato genotype showing differential reaction to salt stress. For example, in the first experiment, 'Siozawa' (V2) showed greater adjustment to the salt treatment via exhibiting a better balance of root and shoot dry matter biomass under induced salt stress whereas 'Chwerotonglo' (V3) maintained superiority in the biological activity. Similarly, 'Gambaru-Ne 3' exhibited greater ability to adjust to the conditions of higher induced salt stress (Figs. 1 and 2). The interaction elucidated from this data reveals change in ranks in the order of performance as observed in the case of V2, V6 and V8 with V7 maintaining superior biological activity in Experiment 2. Thus, the ability or inability of some tomato genotypes to adjust to the varying salt levels via shifts in the shoot:root dry matter biomass balance is likely to be critical for their superior performance under the stress. Mineral content analysis suggested that Na+ and Cl- were accumulated in the roots, stems and finally the leaves in that order for a majority of the tomato genotypes (Table 3). Both genotypic and salinity effects, along with the interactions thereof, were detectable. In Experiments 1 and 2, tomato genotypes; 'Siozawa' and 'Gambaru Ne-3' demonstrated relatively low accumulation of the two ions (Na+ and Cl-) into their leaves at the higher salinity treatment, respectively. DISCUSSION The differential accumulation and re-distribution of dry matter biomass among the study tomato genotypes with respect to root and shoot tissues, under a range of salt treatments suggested variation in biological activity (Figs. 1 and 2). The extremely reduced plant leaf area (Tables 1 and 2) only conferred little surface for vital photosynthetic activities. Most probably, 'Gambaru-Ne 3' has an extremely efficient system for the re-distribution of photosynthates from the source (shoot) to the sink (root), thus, enabling aggressive root production under salinity (Fig. 2). The survival power of a tomato genotype under salt stress depends much on its ability for greater photosynthetic capacity, successful pollination, fruit set and production of viable seeds for evolutionary perpetuation from one generation to the next (Agong et al., 1997). Nonetheless, the ability of a genotype to resist extreme decrease in dry matter biomass production under salinity remains an important criterion for the assessment of salt tolerance as well. In this study, no expressible genotypic differences, except for salt levels, were noticeable on the basis of green meter value (Table 2), thus strengthening the hypothesis that chlorophyll accumulation is unlikely to be significant in fostering salt tolerance ability in tomato (Cuartero and Fernández-Muñoz, 1999). Probably, the chlorophyll content is only enhanced per unit leaf area via the severe reduction in the leaf area under salt stress (Table 1). From the accumulation of Na+ and Cl-, it is apparent that exclusion as a mechanism, facilitated the salt tolerance ability observed in 'Siozawa' and 'Gambaru Ne-3' (Table 3), principally enabling them to behave unlike the wild relatives (Rush and Epstein, 1976). The salt tolerant wild tomato relatives (Lycopersicon peruvianum, L. pennelli and L. chesmanii) survive salt affected niches via the accumulation of high amounts of sodium in their leaves compared to the salt-sensitive L. esculentum (Rush and Epstein, 1991). In general, as would be expected (Agong et al., 1997), the least tolerant genotypes exhibited extreme growth retardation and dry matter accumulation via the excessive toxic Na+ and Cl- levels in their critical organs like the leaf, thus, rendering them less protected and unable to carry out normal physiological activities. However, categorical classification for salt tolerance using exclusion or inclusion of Na+ and Cl- is extremely difficult given the quantitative overlaps among genotypes with respect to the trait (Table 3), and evidence of continuum of sensitivity existing across the plants (Blum, 1988). This study clearly suggests that 'Siozawa' and 'Gambaru Ne-3' are relatively salt tolerant tomato candidates (Figs. 1 and 2; Tables 1, 2 and 3). However, it would be desirable to involve 'Siozawa' in a series of experiments in order to determine its suitability for increasing tomato production in salt affected land. Nonetheless, the physiological supremacy expressed by 'Chwerotonglo', 'Nyanya' 'Joint' and 'BFNT-R' (Figs. 1 and 2), with respect to dry biomass accumulation under salt stress, cannot be ignored. The simplest signal is that, these genotypes should be subjected to trials as potential rootstocks under moderate salinity given their greater ability for root production at higher salinity. On the other hand, 'Gambaru Ne-3' is definitely a suitable candidate as a salt tolerant tomato rootstock coupled with pathological superiority given its strong tolerance against tobacco mosaic virus, fusarium wilt, bacterial wilt and nematode. Elsewhere, multiple-resistance in open-pollinated processing tomatoes to nematode and Fusarium wilt has been reported (Cuartero et al., 1996). A combination of such multiple pathological and salinity stress tolerance in the tomatoes would definitely be a useful technological and economically viable procedure for the exploitation of the vast salt affected lands. Moreover, the majority of tomato producers are small-scale based farmers with only limited production options (FAO, 1999). Salt tolerance in tomato continues to receive a wide range of scientific concerns (Alarcón et. al., 1994; Adams and Ho, 1995; Bolarían et al., 1995; Cayuela et al., 1996; Copeman et al., 1996; Cramer et al., 1995; Foolad and Chen, 1998, 1999; Monforte et al., 1996; Pérez-Alfozea et al., 1996). However, it is our strong conviction that tangible genetic improvement for salinity stress in tomato is definitely not attainable given the strong genetic negative correlation between the tolerance and yielding of the crop (Asins et al., 1993b). Moreover, diluting the salt tolerance ability via yield improvement, results in degradation of the salt tolerance ability in the subsequent progenies (Rush and Epstein, 1981; Agong et al., 1997). Subsequently, we consider the development of very strong salt tolerant tomato rootstocks for the salt prone areas as an optimal compromise. The rootstocks may be very poor fruit yielders but the key point would be the ability to support the tomato scions with the ultimate goal of profitable tomato production. This study reveals that the ability or inability of some tomato genotypes to adjust to the varying salt levels via shifts in the balance between the shoot:root dry matter biomass as elaborated in the correlation between the two parameters and the inherent biological activity are critical in the performance of the genotype under the salt stress (Fig. 2). Therefore, the evidence built from this report is in full support of selecting plants in situ based on their biological activity. Thus, the correlation between the root and shoot dry matter biomass illustrates the physiological bottleneck underlying the adaptation to salt stress and can be exploited as an effective machinery for the evaluation of salt tolerance in the tomatoes. As would otherwise be expected, using relative value of a single trait alone, for example plant height, leaf area, dry matter shoot or root, would mostly lead to a different categorical classification in terms of tolerance to salinity. Hence, the correlationship between shoot and root dry matter, the balance thereof and total dry matter decreasing ratio seem likely suitable criteria for interpreting tolerance to salinity (Figs. 1 and 2). Elsewhere, character definition for understanding salt tolerance in tomato is well documented (Asins, et al., 1993b). However, the problem of discerning the most sensitive and reliable character for interpreting the salt tolerance ability remains largely unresolved (Asins et al., 1993a; Foolad and Chen, 1999). Our results show that there is need to develop strong tomato rootstock for the exploitation of saline ecological niches and also the dilemma existing in the physiological interpretation of tolerance to salt stress. Understanding the physiological responses in tomato due to salt stress is only scarcely documented (Balibrea et al., 1997). Furthermore, quality and availability of water for crop production are recorded as major constraints worldwide (Maas, 1986). Therefore, water economics, and physiological responses to salt stress in tomato genotypes with focus on fruit quality enhancement must be closely investigated into greater depths as we advance into the tomato rootstocks development. The tomato rootstock technology, once developed, will certainly benefit the African farmer in the realisation of fuller utilization the arid land available in the region. ACKNOWLEDGEMENT Thanks to Matsumae International Foundation for granting the fellowship, and to Prof. Dr. M. Masuda for providing the research facilities. Lastly, to Prof. Dr. Stephen Gaya Agong, through whom it was possible to carry out the study. REFERENCES
©2003, African Crop Science Society The following images related to this document are available:Photo images[cs03016t3.jpg] [cs03016t1.jpg] [cs03016t2.jpg] [cs03016f2.jpg] [cs03016f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}