 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
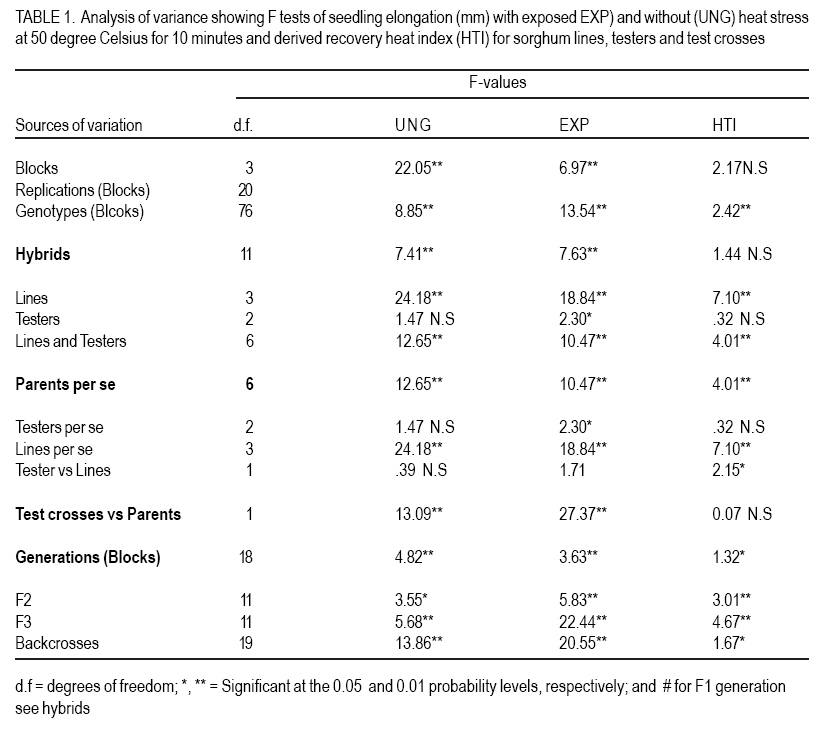
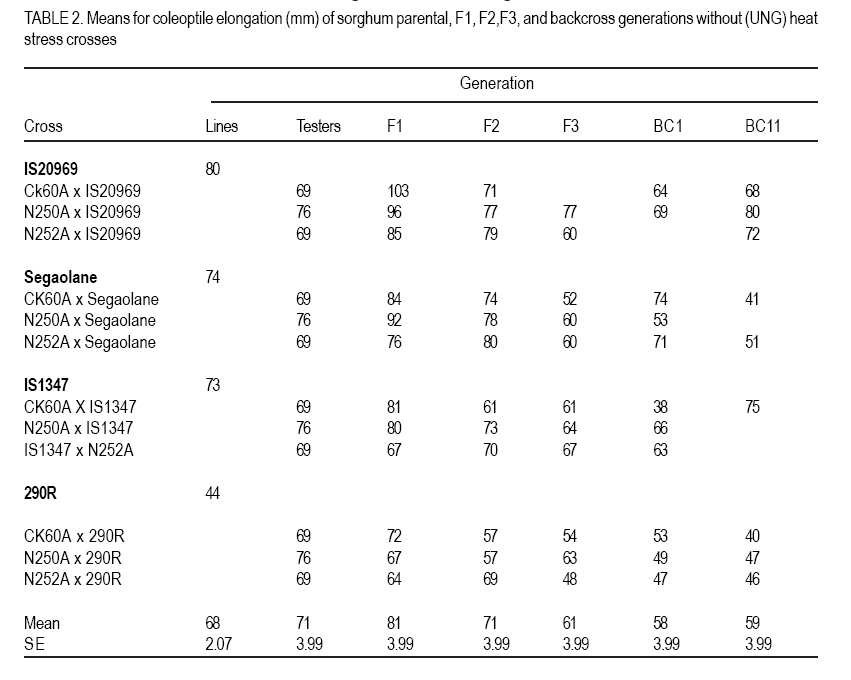
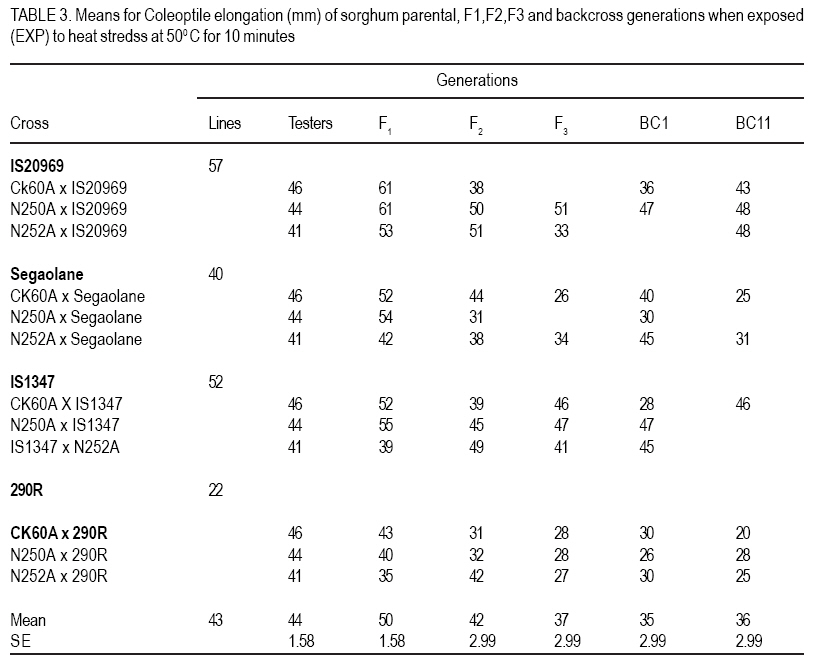
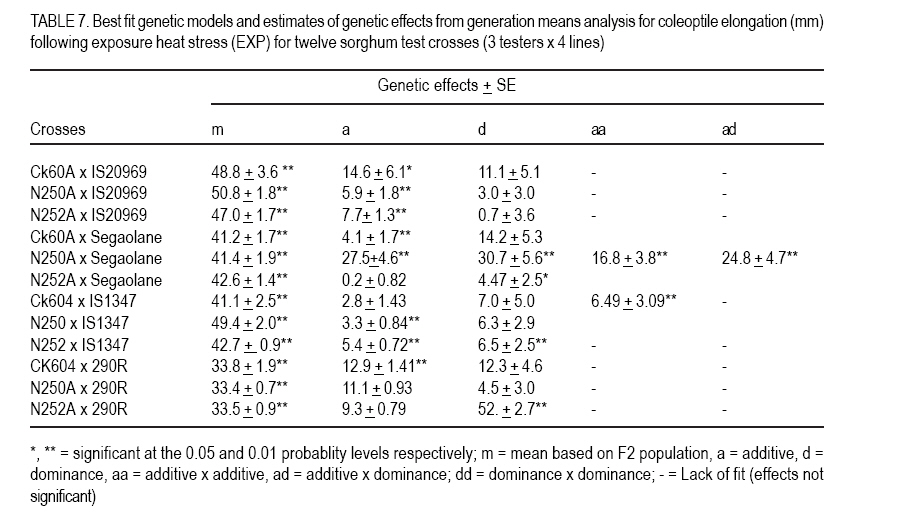
African Crop Science Journal, Vol. 15, No. 1, March, 2007, pp. 33-42 Genetic Evaluation of Seedling Heat Tolerance in Sorghum P.S. Setimela, D.J. Andrews1, K.M. Eskridge1 and J. Partridge1 CIMMYT- Zimbabwe, P.O. Box MP 163, Harare, Zimbabwe Received 9 December, 2006; accepted 1 March, 2007 Code Number: cs07004 ABSTRACT Surface temperatures of tropical soils at planting time, where sorghum (Sorghum bicolor) is a traditional crop, can exceed 50 oC for hours. Seedling heat tolerance is critical for adequate crop establishment in the semi-arid tropics. Improvement of seedlings heat tolerant genotypes would reduce crop losses due to sufficient plant populations. The objectives of this study were to estimate seedling tolerance to heat, determine individual parental contribution and estimate additive, dominance and epistatic effects for seedling tolerance. In our experiments, seedling heat tolerance termed heat tolerance index (HTI) was defined as a ratio of resumed coleoptile growth after a controlled heat shock, compared to normal growth. Genetic parameters of HTI were determined by crossing four lines with varying HTI, with three tester lines, and deriving F1, F2, F3, BC1 and BC11 families for generation means analysis. Line IS20969 from Egypt showed the highest HTI of 0.71, while 290R, an experimental line from the University of Nebraska was the lowest at 0.51. Additive and dominance effects contributed to coleoptile elongation under normal conditions, but only additive effects were significant in recovery growth. Epistatic effects were present in both conditions. General combining ability (GCA) effects for HTI were highly significant in both conditions, but specific combining ability effects were negligible. These results indicate that it is possible to improve seedling heat tolerance and, thus, improve sorghum variety and hybrid plant populations in tropical areas where hot soil temperatures occur. Key Words: Combining ability, genetic effects, heat tolerance index, sorghum bicolor RÉSUMÉ Les températures en surface des sols tropicaux au moment des semis, où le sorgho est une récolte traditionnelle, peuvent excéder 50oC pendant des heures. La tolérance de la chaleur des plantes est critique pour l’établissement adéquat de la culture dans les tropiques semi-arides. L’amélioration des génotypes résistants à la chaleur de plantes réduirait des pertes de récolte dues aux populations de plantes suffisantes. L’objectif de cette étude était d’estimer la tolérance des plantes a la chaleur, déterminer l’additif parental individuel de contribution et d’évaluation, la dominance et les effets épistatiques pour la tolérance des plantes en utilisant des analyses de moyens de génération des populations développées à partir de l’ensemble de détail de parents. Dans nos expériences, la tolérance de la chaleur des plantes l’index nommé de tolérance de la chaleur (HTI) a été définie comme rapport de croissance reprise de coléoptile après un choc commandé de la chaleur, comparé à la croissance normale. Des paramètres génétiques de HTI ont été déterminés en croisant quatre lignes de différents HTI avec trois lignes d’appareil de contrôle, et dérivant les familles F1, F2, F3, BC1 et BC11 pour la génération veut dire l’analyse. La ligne IS20969 d’Egypte a montré le plus haut HTI de 0.71 tandis que 290R, une ligne expérimentale de l’université du Nébraska, était la plus basse à 0.51. L’additif et les effets de dominance ont contribué à l’élongation de coléoptile dans des conditions normales, mais uniquement les effets additifs étaient significatifs dans la croissance de rétablissement. Les effets épistatiques étaient présents dans les deux conditions. Les effets de combinaison généraux de la capacité (GCA) pour HTI étaient fortement significatifs dans les deux conditions mais les effets de combinaison spécifiques de capacité étaient négligeables. Ces résultats indiquent qu’il est possible d’améliorer la tolérance de la chaleur pour des plantes améliores et d’améliorer ainsi la variété de sorgho et les populations de plantes hybrides dans des secteurs tropicaux avec de fortes températures de sol. Mots Clés: Combining ability, genetic effects, heat tolerance index, sorghum bicolor INTRODUCTION Sorghum (Sorghum bicolor L. Moench) evolved in the semi-arid tropics, which are characterised by low rainfall, low soil fertility and high temperatures. Despite the level of its adaptation to the semi-arid tropics, seedling establishment is still a major problem. Failure of seedling establishment due to heat stress is one of the key factors that limit yields and affect stability of production in the semi-arid tropics (Peacock, 1982). According to Radford et al. (1989), only 55% of sorghum seed planted in the field in Australia resulted in successful plants. They also estimated that in Australia, sorghum growers lose 30% of the potential yields due to inadequate plant populations. Uniformity in the establishment of crop stands is one of the essential factors to obtaining a full yield under any production system. A wide range of genetic attributes for thermal tolerance has been reported in many crop species (Levitt,1980). Thomas and Miller (1979) reported that sorghum seedlings respond differently when exposed to varying temperatures, and genetic variation for thermal tolerance in sorghum has been shown to exist in certain lines that are cable of emerging at soil temperature of about 55 oC (Wilson et al., 1982) Beside sorghum, Allan et al. (1962) observed varietal differences in wheat with one variety emerging more evenly in some fields than other varieties under high temperatures conditions. Peacock et al. (1993) and Howarth (1989) have discussed the need for greater diversity in sorghum seedling tolerance to heat in superior genotypes, as this will improve crop establishment in the semi-arid tropics. Genotypes with heat tolerance can be developed as it has been done for other traits such as diseases and insect resistance (Miller, 1982). The objectives of the study reported herein were to gather information on the expression of heterosis of seedling heat stress, understand the contribution of individual parental lines and to estimate additive, dominance and epistatic effects for seedling heat tolerance by utilising generation means analyses using populations developed from a specific set of parents. MATERIALS AND METHODS Experimental materials. This study was based on four genetically diverse sorghum lines and 3 testers (inbred lines used to test the value of genotypes based on performance of their offspring). The genotypes were chosen on the basis of their variable response to heat stress. Three of the parental lines, are known sources of, heat tolerance; IS20969 a late maturing resistant line from Kenya; Segaolane of intermediate maturity from Botswana; IS1347 an early Egyptian maturing line; 290R was obtained from the University of Nebraska (UNL) experimental line susceptible to heat stress. Three testers were used, namely, CK60A & B a previously widely used Texas seed parent with moderate heat tolerance, N250A&B and N252A&B, two genetically different recent seed parent releases from UNL breeding programme of unknown heat tolerance. Each parent line was crossed to each tester, producing 12 F1 hybrids; from each F1 the following generations were developed: BC1 (backcross to parent line only), BC11 (2nd backcross to parent line), F2 and F3. The 12 F1 hybrids were produced using tester cytoplasmic male sterility A-lines but F1 s for deriving progeny were made by plastic bag emasculation on the male fertile line, as were the BC1 and BC11 x F1 s. Because this method does not ensure complete crossing, hybrid plants were identified in each F1 row. Laboratory techniques. Seeds were first cleaned and a uniform median seed size of each genotype was obtained by sieving, which reduced variation within genotypes for seed size. Seeds were surface sterilised by immersion in a hypochlorite solution (1% available chlorine) for 5 minutes and then rinsed thoroughly with distilled water. Fifty seeds from each genotype were placed on a germination paper, moistened with distilled water, and germinated in an incubator for 40 h at 30°C. Germinated seeds (defined to have occurred after 1 mm protrusion of the radicle) were exposed to heat shock using standard procedures and methods described by Setimela et al. (2004). One set from a replication of a generation block was subjected to a direct heat shock by placing the seedlings in a controlled temperature water bath at 50 oC for 10 minutes. The other set of seedlings of the same genotypes were treated similarly but without any heat shock. Prior to treatment, photocopies were taken of all plates to record coleoptile length to derive plot means. Following the treatments the seedlings were returned to the 30oC growth chamber. After 36 h, the length was measured, by photocopying the numbered plates and obtaining the lengths of 10 randomly selected coleoptiles. Statistical design and analysis. The experiment was performed as replication-within block design due to larger number of entries following a procedure outlined by Comstock and Robinson (1948) and Ross and Gardner (1983). The procedure takes each generation as a block. Each block had 5 replications and the same set of generation. This design allowed the comparison of genotypes within the same generation. The entries per set and the sets per generation remained together in the second replication but were again randomised. Each generation experiment can be treated as a small randomised complete block design, involving a set of generations. The parents were included in each generation experiment and the overall mean of the parents in a replication was subtracted from all the entries in that replication were used to remove the variation among generations so that entries are not confounded within blocks. Based on the adjusted values the experiment was reanalysed as a randomised complete block design. Data were subjected to analysis of variance to account for differences among generations. General and specific combining abilities were estimated and tested based on adjusted means. Estimates of genetic effects for each cross were determined by subjecting all populations to generation means analysed by procedure out lined by Hallauer and Miranda (1988). Least squares regression techniques were used on adjusted mean to obtain estimates of parameters: m, a, d, aa, ad and dd which, respectively, represent means based on the F2 additive effects, dominance effects, additive x additive effects, additive x dominance effects, and dominance x dominance effects. Heat tolerance per se was expressed by heat tolerance index (HTI) defined as a ratio of resumed coleoptile growth after the controlled heat shock (exposure time 10 min at 50°C) compared to normal growth. RESULTS AND DISCUSSION Significant differences were detected among generations in the analysis of variance for coleoptile elongation under normal growth (UNG), after heat exposure (EXP) and for the derived heat tolerance index (HTI) (Table 1). The HTI values of lines, F1 and derived generations are listed in Table 2. There were highly significant (P<0.05) differences among genotypes under UNG, EXP and for HTI. There were significant differences between lines and testers in crosses under normal growth and controlled heat shock, but lines vs. testers were only significant for HTI. Similarly, there was remarkable difference between testcrosses and parental lines under heat shock and normal growth. However, testers did not contribute significantly(P>0.05) to differences in heat recovery indices, only CK60A contributed significantly to increased coleoptile growth under normal conditions and under controlled heat shock in its hybrids. Hybrids in general, had higher UNG and EXP values than testers or parental lines, which is an expression of heterosis (Tables 2 and 3). The average UNG of IS20969 test crosses was significantly (P<0.05) better than IS20969, which had the best coleoptile elongation under UNG, EXP and HTI values of any parent (Tables 2, 3 and 4). However, because the EXP of IS20969 test crosses was not better than the parent per se, their HTI were in fact significantly worse than of the higher parent. Segaolane testcrosses though had average EXP and HTI, and were significantly better in these parameters than line itself because Segaolane had below average EXP and HTI values. Though IS1347 did not produce any significant differences, it numerically gave the best HTI with CK60A and N250A and was average with N252A. The experimental line (290R) had low UNG and EXP values; though the UNG and EXP of all 290R testcrosses were the lowest, they were significantly better than 290R per se their HTI showed no change. There were highly significant GCA effects for specific lines with and without heat stress. The GCA effects were consistent among lines with and without heat stress (Table 5). Lines IS20969 and 290R combined well with testers for UNG by highly significant GCA toward the hybrids. The HTI was significant for 290R, but not significant for all other lines. In terms of GCA, IS20969 and 290R were the only parents, which had highly significant estimate (P=0.001) for GCA for coleoptile elongation for UNG and EXP (Table 5). The estimate for GCA for IS20969 was negative, while that of 290R was positive. The signs are expected between the two parents since they have varying degree of tolerance to heat stress. Heterotic effects, however, were in both positive and negative directions with respect to high and low parents. The SCA estimates are ultimately important to test the performance of parents in hybrid combinations. The best parents could be used to produce hybrids. In the present study none of the crosses had significant SCA effects for coleoptile elongation for UNG and EXP. Heterosis in the F1 generation was mostly due to GCA. An additive-dominance model fits the data in most of the crosses. The data fitted a more complex model (epistasis) in two crosses involving N250A x IS20969 for UNG and N250A x Segaolane for EXP conditions (Table 6 and 7). Five of the crosses, had significantly negative additive effects except for a cross involving CK60A x Segaolane under normal conditions. Six of the crosses had a positive significant dominance effect (Table 6). The magnitude of dominance was higher than the additive effects in most of the crosses. Additive x additive epitasis gene effects were significant only in two crosses. Most of the crosses involving the parental line IS20969 had positive significant additive effects and crosses involving 290R had negative significant additive gene effects. The above results indicate that under normal growth both dominant and additive gene effects play a major role for coleoptile elongation. Contrary to normal growth under controlled shock, 10 of the 12 crosses had a significant additive gene effect and only 4 of the crosses had a significant dominance gene effect (Table 7). Two of the crosses involving IS20969 contributed positively to coleoptile elongation under controlled heat shock and all the crosses involving 290R have significantly negative additive effects. The contribution between the 2 parental lines is expected due to their different levels of heat tolerance. The contribution of epistasis to coleoptile length under heat stress was significant only in 1 cross (N250A x Segaolane) indicating that for this particular cross, inheritance of heat tolerance is complex. The additive x additive gene interaction was positive while additive x dominance gene interaction was negative. The epistasis gene with positive and negative effects may have a negative impact on coleoptile elongation as the effects cancel each other (Mather and Jinks, 1982). The performance may depend on the magnitude of the epistasis of the cross involved. With reference to derived HTI, eight crosses had significant additive gene effects and only 1 cross had significantly negative dominant effects (Table 8). The crosses involving IS20969 had significantly positive additive effects and crosses involving 290R had a significantly negative additive gene effect. The predominance of additive gene action for coleoptile elongation under controlled heat shock suggests that improvement can be achieved by selecting in the early generations. The lack of significant SCA for coleoptile elongation indicates that under heat stress; additive gene effects mostly influence elongation. Analysis of variance of the crosses among lines indicated that most of the variation within each set of crosses was attributable to GCA of the lines included in the crosses. These results indicate that when parents have poor coleoptile elongation, significant heterosis manifests, and when coleoptile elongation is intermediate, negative heterosis is observed. Crosses that have lower decline in heterosis across generations indicate less inbreeding depression. CONCLUSION In this study the focus was on seedling tolerance or sensitivity to heat and does not imply resistance or sensitivity to any other stage, although the genotypes were selected based on the level of tolerance or sensitivity on mature plants in the field. Heat tolerant lines did not contribute significantly to hybrids with heat tolerant testers but heat tolerant lines contributed positively to hybrid performance when poor testers were used as pollinators. Dominance effects in most of the crosses were not significant genetic effects were predominant in coleoptile for growth after heat exposure, which explained elongation recovery under heat stress in most of the lack of high heterosis in the F1 generation. the crosses. Epistasis was important only in a These results indicate that selecting potential heat few crosses. Individual genetic effects in some tolerant lines for hybrid production should crosses were positive and negative which improve female parents with poor HTI. Additive indicates the diversity in the genotypes used. The method used to detect significant differences between parents, and the type of gene action involved in this study are useful to a plant breeder to reduce the number of entries before embarking on a field trial. For improvement of coleoptile elongation under heat stress the breeder should use selection procedures emphasizing GCA. REFERENCES
©2007, African Crop Science Society The following images related to this document are available:Photo images[cs07004t4.jpg] [cs07004t1.jpg] [cs07004t7.jpg] [cs07004t8.jpg] [cs07004t6.jpg] [cs07004t5.jpg] [cs07004t2.jpg] [cs07004t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}