 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 15, No. 2, June, 2007, pp. 73-81 Response of Pigeonpea Genotypes of Different Maturity Duration to Temperature and Photoperiod in Kenya S.N. Silim, E.T. Gwataa, R. Coeb1 and P.A. Omanga2 International Crops Research Institute for the Semi-Arid Tropica (ICRISAT), P. O. Box 39063,
Nairobi 00623, Kenya Received 1 October, 2004; 3 March, 2007 Code Number: cs07009 ABSTRACT Pigeonpea (Cajanus cajan (L.) Millsp.) is one of the major grain legumes grown in the tropics and subtropics. The crop is grown rainfed in prone drought areas where day length varies from 11 to 14 h and large differences in temperature are experienced, largely due to variations in altitude and latitude. Field studies were conducted with different pigeonpea [Cajanus cajan (L.) Millsp.] in Kenya to determine the effect of photoperiod and temperature on flowering. Variation in temperature was achieved by planting six genotypes at four locations varying in altitude where temperature decreased with increase in altitude and variation in photoperiod was achieved through artificial lighting (about 12.6 hr - natural day length, 14.5 hr and 16.0 hr). The genotypes used in the study were carefully selected to represent different maturity duration (extra-short-, short-, medium- and long maturity duration) and major piegonpea production regions. Equations that describe the rates of development (1/f) were used to determine rates of progress of each genotype towards flowering as influenced by temperature and photoperiod. For photoperiods below 13 hr, rates of progress towards flowering were influenced by temperature in give genotypes (ICPL 90011, ICPL 87091, ICP 7035, ICP 6927 and ICEAP 00040). The optimum temperature for rapid flowering were 24.7 oC for the extra-short-duration genotype, 23.1 oC for the shortduration genotye, 23.8 and 22.2 oC for medium-duration genotypes and 18.3 oC for the long-duration genotypes, 22.2 oC for medium-duration genotypoes and 18.3 oC for the long-duration genotypes which indicated that the area of origin had a strong influence on adaptation. The effects of photoperiod on rates of progress towards flowering were investigated only under sub-optimal temperatures. The extra-short-duration genotype (ICPL 90011) was the least responsive to variation in photoperiod, while the two long duration genotypes (ICEAP 00040 and T-7) were to most sensitive to photoperiod variation with flowering rate reduced by 0.001 d-1 per hour increase in day length. Key Words: Cajanus cajan, day length, flowering RÉSUMÉ Le pois pigeon (Cajanus cajan (L.) Millsp) est une des principales légumes plantées dans les régions tropicales et subtropicales. La culture est alimentée par les pluies pour sa croissance dans les zones susceptibles de sécheresse, au sein desquelles la durée de la journée varie de 11à 14h et des grandes différences de température sont observées, en majorité dues aux variations d’altitude et de latitude. Des études de terrain étaient conduites avec différents types de pois pigeon [Cajanus Cajan (L.) Millsp.] au Kenya en vue de pouvoir déterminer l’effet de la photopériode et de la température sur la floraison. Les variations de température étaient réalisées au moyen de six génotypes plantés dans quatre endroits d’attitude différentes où la température baissait en fonction d’un accroissement en attitude tandis que la variation de photopériode était atteinte par éclairage artificiel (Environ 12,6 hr – longueur de journée naturelle, 14, 5hr et 16, 0hr). Les génotypes utilisés au cours de l’étude étaient attentivement choisis en vue de pouvoir représenter des durées de maturité variables (extra-courte, courte, moyenne et longue) ainsi que les principales régions productrices de pois pigeon. Des équations décrivant le taux de développement (1/f) étaient utilisées dans le but de déterminer le taux de progression de chaque génotype vers la floraison tel qu’influencé par la température et la photopériode. Pour des photopériodes ne dépassant pas 13 hr, ces taux étaient influencés par la température dans des génotypes donnés (ICPL 90011, ICPL 87091, ICP 7035, ICP 6927 et ICEAP 00040). La température optimale pour une croissance rapide étaient de 24.7oC pour le génotype à durée extra-courte, 23,1oC pour celui à durée courte, 23,8oC et 22,2oC pour le génotype à durée moyenne et enfin 18,3oC pour celui à longue durée; ce qui montrait que la région d’origine présentait une forte influence sur l’adaptation. Les effets de la photopériode sur le taux de progrès vers la floraison étaient recherchés seulement sous températures sub-optimales. Le génotype à durée extra-courte (ICPL 90011) était le plus réceptif quant aux variations de photopériode, pendant que les 2 génotypes à durée longue (ICEAP 00040 et T-T) étaient les plus réceptifs à la variation de la photopériode, avec des taux de floraison réduits de 0.0001 d-1 par heure d’augmentation en longueur de journée Mots Clés: Cajanus cajan, longueur de journée, floraison INTRODUCTION Pigeonpea (Cajanus cajan (L.) Millsp.) is one of the major grain legumes grown in the tropics and subtropics. The crop is grown rainfed in prone drought areas where day length varies from 11 to 14 h and large differences in temperature are experienced, largely due to variations in altitude and latitude. The traditional varieties grown are mainly medium-and long-duration types. They are intercropped with cereals such as maize (Zea mays) and sorghum (Sorghum bicolor) and various short-duration legumes such as cowpea (Vigna unguiculata) and beans (Phaseoulus vulgaris) (Silim et al., 1991). These traditional genotypes are extremely sensitive to photoperiod and temperature, with plant height, vegetative biomass, phenology and grain yield being the traits most affected (Whiteman et al., 1985). The vegetative phase of development normally coincides with relatively long days and warm temperatures at the higher latitudes (Reddy and Virmani, 1981) and with warm temperatures at the higher latitudes (Reddy and Virmani, 1981) and with warm temperatures at lower latitudes (Omanga et al., 1995). Reproductive phase of development coincides with the period when temperatures are cool and day- length is short and as a result, there is a tendency to conclude that flowering is triggered only by short days, thus overlooking the effect of temperature. The sensitivity of pigeonpea to temperature and photoperiod is a major constraint to development of stable and predictable management practices, cropping systems, and genotypes (Whiteman et al., 1985). If sensitivity in phenology leads to a delay in maturity, it is likely to result in yield reduction in those areas where rainfall duration is short or where the crop depends on residual soil moisture. In areas with a bimodal rainfall, such as Kenya, acceleration of phenology may result in reproductive growth occurring between the two rainfall periods and the crop would thus suffer from drought stress at the time when it is most sensitive. Similarly, where there is accelerated phenology, reproductive growth may coincide with a period of high pest incidence (Wallis et al., 1981). Extra-short and short-duration pigeonpea genotypes developed by the International Crops Research Institute for the Semi-Arid Tropics (ICRISAT), escape drought and are relatively less sensitive to photoperiod than traditional varieties with longer growth cycles (Singh et al., 1990). This has increased the flexibility of pigeonpea cultivation and facilitated its use in different cropping systems (Nene, 1991). However, these new genotypoes developed by ICRISAT are sensitive to temperature and are not adapted to cool production areas (Simim and Omanga, 2001). Richards (1989) reported that the first step towards maximising yield is to ensure that the phenology of the crop matches the resources of the production environment. Summerfield et al. (1991) described a series of photothermal models used to predict phenological events such as time to flowering (f) as well as the rate of progress towards flowering (l/f). Such models are useful in selecting prospective genotypes for production in environments that have optimum production conditions. The temperature range in which plant growth and development occurs is characterised by base temperature (Tb) below which the rate of development is zero, an optimum temperature (To) at which the rate of development is most rapid, and a warmer ceiling limit (Tce) beyond which development ceases (Summerfield et al., 1991). In short-day plants such as pigeonpea, flowering response to photoperiod is determined by the critical photoperiod (Pc), defined as that day length which, if exceeded, flowering is delayed. With further increase in daylength, a ceiling photoperiod (Pce)is reached when maximum number of days is taken to reach flowering (Keatinge et al., 1996). There is lack of information on the sensitivity of pigeonpea to temperature and photoperiod under field conditions in the agro-ecological areas in Kenya. Therefore, the objectives of this study were to determine:
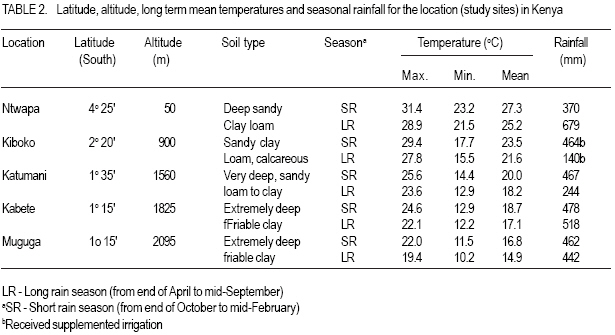
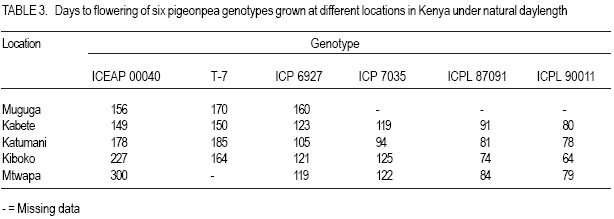
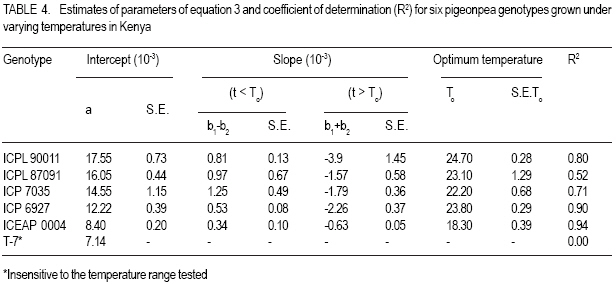
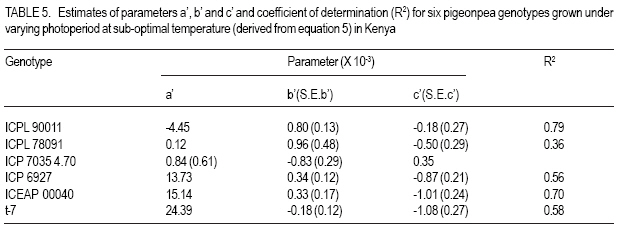
MATERIALS AND METHODS Genetic material. Six genotypes were selected carefully to represent the main maturity duration types found in pigeonpea (Table 1). The extrashort-and short-duration genotypes were developed at ICRISAT. They were selected, as representative genotypes for further evaluation, from the study reported by Silim and Omanga (2001) involving 64 extra short- and shortduration pigeonpea genotypes. The remaining genotypes were selected from 48 genotypes used in the studies reported by Silim et al. (1995). The two medium-duration genotypes (ICP 7035 and ICP 6927) originated from contrasting environments (Table 1). ICP 7035 was a selection from a landrace collected in India, while ICP 6927 was a collection from Trinidad and Tobago. Similarly, the two long-duration genotypes were from contrasting environments, with T-7 originating from northern India but ICEAP 00040 originating from Kenya. Environments. The experiments were conducted at five locations in Kenya, varying in altitude from 50 m to about 2100 m, above sea level and where temperatures decrease with increase in altitude (Table 2). The experimental sites covered traditional (Katumani, and Kiboko) and nontraditional (Mtwapa, Kabete and Muguga) pigeonpea growing areas. At the experimental sites, latitude ranged from 4o 25' to 1o 15' S and the daylength was about 12.6 hr. Experimental design. There were three light treatments, natural daylength (about 12.6 hr) in all locations, 14.5 hr (Kiboko, Katumani and Kabete) and 16 hr (Kiboko and Kabeter). Exposure of genotypes to longer daylengths was achieved by use of 100 W incandescent bulbs that were suspended 2.0 m above ground and 1.5 m apart. The 14.5 hr daylenght was achieved through an automatic switch that turned the lights on at 05 hr and off at 07 hr in the morning and on in the evening at 18 hr and off at 19.5 hr. The 16 hr day length was achieved by turning the lights on at 05 hr and off at 07 hr in the morning and on in the evening at 18 hr and off at 21 hr. The daylength treatments were sited at least 100 m from each other. Each daylength treatment was designed as a separate trial. Each trial was conducted using six pigeon genotypes planted in a randomized complete block design and replicated three times. At each site, the first planting (for the long rain season) and second planting (for the short rain season) were performed (at the start of the rain seasons) during the first week of April and November, respectively. Each genotypes was sown in plots consisting of three 5 m rows. Extrashort-and short-duration genotypes were planted at 0.4 m between rows and 0.1 within rows. Medium duration genotypes were planted at 1.0 m between rows and 0.5 m within rows. Longduration genotypes were planted at 1.2 between rows and 0.5 within rows. Sowing was by hand in ploughed and harrowed fields. Three to four seeds were sown in each hill. During planting, no fertilisers were used, neither were the seeds inoculated with rhizobia. Fourteen days after emergence seedlings were thinned to one per hill. Weeds were controlled by hand weeding. The plants were protected against insect pests during flowering and post-flowering phases by insecticidal sprays. No mineral deficiency symptoms were observed during crop growth. At Kiboko, the driest location, the trials were maintained as near as possible to field capacity through supplement irrigation. At the other locations, the crop did not experience water deficits. Data collection and analysis. During the duration of the trials, daily maximum and minimum temperatures were obtained from meteorological stations located within 400 m of all of the experiments. The mean pre-flowering maximum (Tmax.) and minimum (Tmin.) temperatures for each genotype in each environment were calculated using the daily temperature values from sowing until 50% of the plants in a plot had at least one open flower (flowering). Time to flowering was recorded for each genotype. The mean pre-flowering photoperiod for each genotype was calculated from the daily photoperiod between sowing and flowering. The response to temperature and photoperiod of times from sowing to flowering (f) was based on means of three replications for each genotype-environment. Rates of progress from sowing to flowering were calculated as 1/f (i.e. the reciprocal of the time from sowing to flowering) for each genotype using the protocol developed by Summerfield et al. (1991, 1993). The protocol is based on three well-defined linear relations between 1/f and mean pre-flowering temperature (T) and mean pre-flowering photoperiod (P). In a crop such as pigeonpea, which is photoperiod sensitive, when daylength is less than critical photoperiod (Pc), then 1/f is a positive and linear function of temperature from a base temperature (Tb) at which the rate is zero, up to an optimum temperature (To) at which it is maximal. Between Tb and T o, the relationship is described as: 1/f = a + bT ............................................ Equation 1 where the values a and b are specific to each genotype; above To the values of the constants are different from those in the sub-optimal range and relation (i.e., the sign of the parameter b) will be negative. Depending on the photothermal environment for a given duration type, additional procedures described by Omanga et al. (1995) were applied. In these procedures, the data for each genotype were fitted to: 1/f = a1 + b1 (T - To) + b2 IT-T oI ........... Equation 2 where To is the apparent optimal temperature to be estimated, a1 is the apparent maximal rate of progress to flowering when T = To, the parameter constants b1 and b2 combine to determine the flowering response to the entire temperature range from Tb (base) to Tc (ceiling) and the symbol 1 1 is the absolute value of the difference between T and To. The above equation can be rewritten as: 1/f = a1 + (b1 - b2) (T - To) for T< To 1/f = a1 = (b1 + b2) (T -To) for T > To .... Equation 3 where a1, b1 and b2 are linear parameters to estimate. To requires non-linear estimation. In short day plants, there is critical photoperiod (Pc) which is the daylength when exceeded, causes a delay in flowering, and a ceiling photoperiod (Pce), longer than Pc when f reaches a maximum number of days (Summerfield et al., 1991)? The rate of development a given temperature within the limits of the Pc and Pce is given by: 1/f = a’ + C’P ........................................ Equation 4 where P is daylength (h d-1) and a’ and c’ are genotype specific constants of which c’ has a negative value for a short-day plant such as pigeonpea. In addition, a maximum delay is reached at the Pce above which there is no further delay in flowering in short-day plants and when P ≥ P ce, a variation in P or T has no effect on f. In most crops, both temperature and photoperiod modulate progress towards flowering. A combines equation describing variation in rate (1/f) in photoperiod-sensitive genotypes between critical and ceiling photoperiods and between the base and optimal temperatures may be described as a response plane give by: 1/f = a’ + b’T + c’P ............................... Equation 5 and the plane of maximum photoperiod delay in given by: 1/f = d’ .................................................. Equation 6 where a’, b’, c’ and d’ are genotype specific constants. RESULTS Mean temperatures experienced in each of the ten environments created by locations and seasons indicated that the plants were exposed to different temperatures during both pre- and post-flowering development phases (Table 2). The post-flowering temperatures were cooler than the pre-flowering temperature during the long rain season (for the crop sown in April), but warmer during the short rainy season (for the crop sown in November). Medium - and longduration genotypes experienced cooler temperatures during the reproductive phase at Kiboko, Katumani, Kabete and Muguga (Table 2). There were large variations across the environments for mean f (Table 3). Both the extrashort and short-duration genotypes flowered within 100 days from sowing but the medium and long - duration genotypes flowered later. For photoperiod < 13 h, rates of progress towards flowering in five genotypes (ICPL 90011, ICPL 87091, ICP 7035, ICP 6927 and ICEAP 00040) were affected by temperature (Table 4). The extra-short duration genotype (ICPL 90011) had the highest optimum temperature while the long-duration genotype ICEAP 00040 had the lowest optimum temperature. The long duration genotype (T &), which originated from northern India (where temperatures vary from sub-zero to > 40 oC during the growing season), was insensitive to temperature in the range tested. There was no clear pattern in the slopes of the response of different genotypes to temperature (Fig. 1). However, at sub-optimal temperatures, the genotypes originating from warm environments (ICPL 90011, ICPL 87091 and ICP 7035) relatively, were more sensitive to temperature in the range tested. There was insufficient data at the supraoptimal temperature range to develop models for response to photoperiod under the range suboptimal to supra-optimal temperature. As a result, response to photoperiod was analysed using the model in equation (4) fitted to data from suboptimal temperatures only (Table 5). The genotype ICPL 90011 was insensitive to daylength changes but the long-duration genotypes (ICEAP 00040 and T - 7) showed a strong response, with flowering rate reduced by 0.001 d-1 per hour increase in daylength. DISCUSSION The six pigeonpea genotypes used in this study were selected to represent the four duration types (extra-short, short-, medium- and long-duration) as per ICRISAT classification and the major pigeonpea agro-ecologies in Kenya. The extra-short- and short-duration genotypes had high optimum temperature for time to flower, with cool temperatures lengthening and warm temperatures shortening the duration. This observation was consistent with findings reported in similar studies (McPherson et al., 1985; Turnbull and Ellis 1987; Silim and Omanga 2001). For medium-duration genotypes, the differences in optimum temperature for time to flower between the two genotypes indicated their adaptation to area of origin. ICP 6927 originated from lowland areas near the equator where temperatures are high and with low seasonal variability; while ICP 7035 is from low elevation areas where temperatures during the vegetative stages are high but low during the reproductive phases. By contrast, the long-duration genotype (T - 7) was insensitive to variation in temperature likely because it is a landrace from northern India where temperatures during the crop’s growth cycle vary from >40 oC to sub-zero. The result from this study also indicated that adaptation for pigeonpea can be classified into three broad categories, namely (i) genotypes bred in (or evolved from) areas with high temperatures (e.g. ICPL 90011, ICPL 87091, ICP 6927) have high optimum temperature for f (with low temperatures delaying f), (ii) genotypes from medium altitude areas (e.g. ICEAP 00040) where mean temperatures vary from warm (21 oC) to cool (17 oC), have low optimum temperatures f (with high temperatures delaying or inhibiting f), and (iii) genotypes that originated in areas where variation in temperatures are large (e.g. T - 7) are insensitive to temperature in terms of f. This study also showed that with the exception of genotypes from northern India, pigeonpea has specific and narrow adaptation. In barley, Ceccarelli (1989) suggested breeding either for narrow or wide adaptation in barley and proposed the need for defining the target environment even for wide adaptation. Knowledge of the environment where the genotypes are bred (or originate) would be useful in designing pigeonpea breeding progammes and selecting genotypes for cultivation in specific optimum environments. For photoperiod, the long duration genotypes (ICEAP 00040 and T - 7) were the most sensitive, followed by medium-duration genotypes (IVCP 6927 and ICP 7035), which were intermediate in sensitivity. The short-duration genotype (ICPL 87091) was insensitive and extrashort-duration genotype (ICPL 90011) was the least sensitive to photoperiod. Carberry et al. (2001) reported that medium- and long-duration cultivars delayed flowering by 150 days in response to photoperiod. The observed lack of sensitivity in these genotypes could be attributed to their place of origin in India, where planting is carried out during the longest days in June, and flowering occurs at the beginning (for ICPL 90011) and end (for ICPL 87091) of August under long but progressively decreasing days. In most countries in eastern Africa, longduration types of pigeonpea are intercropped with maize. In cool areas, severe competition occurs between intercropped maize and pigeonpeas resulting in yield reduction of both crops. Therefore, we propose the development, through plant breeding, of long-duration cultivars that mature later than the current traditional longduration genotypes. Secondly, current mediumduration genotypes from the equatorial region mature 50 to 100 days earlier than the longduration types. However, because of their sensitivity to photoperiod, the current mediumduration genotypes tend to mature late when grown in areas away from the equator, such as Malawi and thus suffer from terminal drought stress. In our view, it is necessary to introgress insensitivity to photoperiod but maintain high optimum temperatures in these genotypes in order to use the, in areas away from the equator. REFERENCES
The following images related to this document are available:Photo images[cs07009t1.jpg] [cs07009t4.jpg] [cs07009t5.jpg] [cs07009t2.jpg] [cs07009t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}