 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 15, No. 3, Sept, 2007, pp. 127-138 Within Plant Resistance to Water Flow in Tomato and Sweet Melons S.O. Agele and S. Cohen1 Department of Crop, Soil and Pest Management, Federal University of Technology,
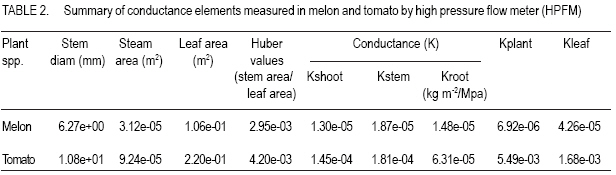
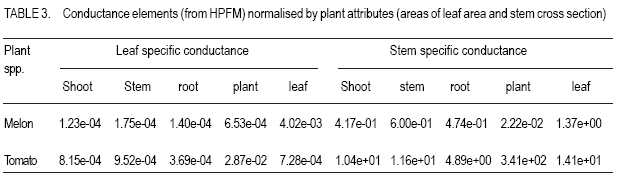
PMB 704, Akure, Nigeria (Received 1 July, 2004; accepted 10 March, 2007) Code Number: cs07014 ABSTRACT Efficient water resource management in relation to water use and crop yields is premised on the knowledge of plant resistance to water flow. However, such studies are limited and for most crops, the within plant resistance to water flow remains largely unknown. In this study, within plant resistance to water transport (hydraulic conductance) was monitored in tomato (Lycopersicum esculuntum) and sweet melon (Citrullus lanatus) using the high pressure flow meter (HPFM) and evaporative flux (EF) methods. In the evaporative flux method, measurements of transpiration flux and leaf water potential were used to calculate the total resistance to water flow using Ohm’s law analogy. Measurements of tranpiration flux (Q) relationship, plant resistance calculated from the slope of their relationship, ranged from 6.57x10-01 to 2.27x10-03 Mpa m-2s-1 for tomato and sweet melon, respectively. The magnitude of whole plant hydraulic conductance calculated by the evaporative flux method and measured on the HPFM were not significantly different. This is probably due to that fact that Kp includes the hydraulic conductance of the root system, which offers the highest resistance to water flow in a plant, and the frictional resistance of the proximal part of the crown. Day time course of water relation parameters were monitored in melon and tomato (predawn, 1100 to 1400 h). The effect of time of day was pronounced on the dynamics of water relations, particularly around mid-day when sharp differences in the magnitudes of the measured water relation parameters were obtained. Minimum (predawn) leaf water potential (ψl) ranged from 0.135 to 0.207 MPa for both crops. The greatest conductance was found in the leaf and the lowest in the root in both crops. While the highest within plant resistance to the flow is contained in the root system, the stem component constitutes the least resistance (greatest conductance) to within plant flow of water in tomato. Repeated measurement analysis showed the existence of significant species effects on plant water relations (leaf water potential, evaporative/transpiration flux, xylem hydraulic conductance). Superiority within plant xylem transport and evaporative losses were obtained in tomato, from the relations of EF and Kh; this crop showed a higher hydraulic sufficiency than melon. The expression of hydraulic conductance of the root and shoot system relative to plant attributes did not eliminate differences in the magnitudes of conductance elements in tomato and melon. Differences obtained between melon and tomato in whole plant leaf and stem area specific hydraulic conductance (KI) indicate the carbon efficiency and, hence, the cost of resource allocation to areas of root surface and leaves for enhanced scavenging for water and mineral nutrients. Thus, application of mineral nutrients to enhance crop productivity should take into consideration the hydraulic conductance for specific plants. Key Words: Citrullus lanatus, evaporative flux, Lycopersicum esculuntum, transpiration RÉSUMÉ La gestion efficace de ressource d’eau en relation avec son utilisation et son rendement est une prémisse de la connaissance de la résistance des plantes au flux d’eau. Cependant, de telles études sont limitées et pour la plupart des récoltes, la résistance interne de la plante sur le flux d’eau reste principalement inconnue. Dans cette étude, la résistance de plantes sur le flux (la conductance hydraulique) a été contrôlée dans la tomate (Lycopersicum esculuntum) et le melon doux (Citrullus lanatus) utilisant la méthode de forte pression (HPFM) et le flux d’évaporation (EF). Dans la méthode de flux d’évaporation, les mesures de flux de transpiration et de potentiel d’eau de feuille ont été utilisées pour calculer la résistance totale en utilisant l’analogie de loi d’Ohm. La relation entre les mesures de flux de transpiration (Q) et la résistance des plantes a été calculé, la pente de cette relation, était entre 6.57x10-1 à 2.27x10-3 MPa m-2s-1 pour la tomate et le melon doux, respectivement. La grandeur de la conductance hydraulique de la plante entière prédite par la méthode de flux d’évaporation et mesuré sur le HPFM n’était pas significativement différente. Ceci est probablement en raison du fait que Kp inclut la conductance hydraulique du système fondamental, qui offre la plus haute résistance au le flux dans une plante, et la résistance de friction de la partie proche de la couronne. L’effet du temps de la journée sur les paramètres d’eau a été contrôlé dans le melon et la tomate (de 11.00 à 14.00 h). L’effet du temps a été prononcé sur la dynamique de relations d’eau, particulièrement autour du midi quand les différences tranchantes en grandeur des paramètres de relation d’eau mesurés ont été obtenues. Le potentiel d’eau minimal de feuille était compris entre 0,135 et 0,207 MPa pour les deux récoltes. La plus grande conductance a été trouvée dans la feuille et le plus bas dans la racine dans les deux récoltes. Pendant que la plus grande la résistance interne de la plante au flux est contenue dans le système fondamental, le composant de tige présentait la moindre résistance (la plus grande conductance) au flux d’eau dans la tomate. L’analyse répétée de mesure a montré l’existence d’effets d’espèce significatifs sur les relations d’eau des plantes (le potentiel d’eau de feuille, le flux de d’évaporation transpiration, la conductance hydraulique du xylem). La supériorité dans le transport de xylem de plante et dans les pertes d’évaporation a été obtenue dans la tomate, concernant EF et de Kh; cette récolte a montré une efficacité hydraulique supérieure au melon. L’expression de la conductance hydraulique de la racine et le système relatif aux attributs de la plante n’a pas éliminé les différences dans les grandeurs de la conductance dans la tomate et le melon. Les différences ont été obtenu entre le melon et la tomate dans la feuille de la plante entière et le secteur de tige pour la conductance hydraulique spécifique (KI) indiquant l’efficacité du carbone et, donc, le coût d’allocation de ressource aux secteurs de surface et de feuilles fondamentales pour améliorer la récupération de l’eau et de nutriments minéraux. Ainsi, l’application de nutriments minéraux pour améliorer la productivité de récolte doit prendre en considération la conductance hydraulique des plantes. Mots Clés: Citrullus lanatus, flux d’évaporation, Lycopersicum esculuntum, transpiration INTRODUCTION The hydraulic transport efficiency of a plant is an important ecophysiological determinant of survival and growth under varying environmental conditions; for example, soil and air drought. Plant resistance is modulated by changes in the status of water conducting system, water transport processes in addition to biological or physiological regulations. Within plant, resistance in the root to leaf water flow pathway is not constant, but increases with decreases in transpiration flux (Steudle, 1994). In addition, whole plant water balance may depend more on the changes in the physical status of the plant water conducting system than on biological regulation of root water permeability (LoGullo et al., 2003). The ability of plants to maintain a favourable water status is dependent on the resistance to water flow in the Soil-Plant-atmosphere coupling (SPAC) (Jones et al., 1982; Ruggiero et al., 2003). Total resistance (Rtotal) in the water transport pathway in the SPAC controls water flow, however, the resistance that most limits plant water uptake is subject to debate (Ruggiero et al., 1999). Under non-limiting soil water status, within plant resistance dominates soil resistance to water flow; however, increases in total resistance in the SPAC may be due to increase in soil resistance (conductivity of the unsaturated soil), or higher resistance in the liquid water flow within the plant than in the soil. Within the plant, root resistance to water flow is predominant with xylem axial resistances smaller than the radial resistance (Tyree and Sperry, 1989), however, soil hydrailic resistance and root densities would be limiting resistance in dry soils (LoGullo et al., 2003). Environmental factors may trigger responses in the components of the total resistance to water flow in crops. Plant hydraulic properties change in response to the internal and external cues and environmental factors (Zhu and Steudle, 1991; Ruggiero et al., 2003). In plants, morphological or physiological adjustments are known to be associated with the maintainance of homeostatis of physiological funtions (Gyenge et al., 2003; Sobrado, 2003; Tyree, 2003). Differences in the transport of water to leaf surfaces and separate control of water use ecophysiological attribute with impact on the performance of a species in contrasting habitats (Gyenge et al., 2003; Sobrado 2003). Hence, high transpiration rates via within plant water transport efficiencies would sustain biomass accumulation and, hence, survival and growth. Therefore, the plasticity of the hydraulic system is useful input in breeding programmes for crop varieties adapted for cultivation under varying environmental conditions (Mencuccini, 2003; Tyree, 2003). Soil-Plant-atmosphere coupling (SPAC) explain the control exercised by the soil and the atmospheric conditions (environments) on plant processes. Literature shows that changes in plant water relation parameters like leaf water status and stomatal response can be explained in terms of changes in the hydraulic architecture of plants (Zimmermann, 1983; Tyree and Cochard, 1996; Saslleo et al., 2000). Extensive studies have been reported for woody species (trees) on the role/mechanisms of hydraulics and stomatal conductances in the soil-plant-atmosphere continuum (Ruggiero et al., 1999; Tsuda and Tyree, 1999; LoGullo et al., 2003); however only a few studies have addressed these issues in annual crops (Ruggiero et al., 1999; 2003) and very little is known about tropical crops under rainfed agriculture. The dependence of the hydraulic property on the water relation of root and leaf and the functional linkage of stomata conductance (gs) of the leaves to the hydraulic conductance of the soil-leaf continum merits further studies among crop species (Sperry, 2000). Among herbaceous species, there is need to quantify components and magnitudes of variations of the resistance to water flow during growth cycle in terms of the effects of such variations or response on the water flow and implications for survival and productivity under varied weather environments of the growing seasons. The understanding of plant water relations in crop species have implications for crop water use, crop survival, functioning and productivity in contrasting habitats/growing environmental conditions, and therefore, to water resources planning and management. Knowledge of water use is required to manage the water status of crops for a desired performance over the season in a given climate and to the maintenance of ecosystem water balance. Since evaporation from the soil and canopies of vegetation and transpiration of crop species (weeds) need to be combined in order to infer whole ecosystem water use, an important component of water resources management. Extrapolation of water consumption by agricultural crops in the humid tropics from published data from other agroecologies may not be valid due to differences in soil and soil water holding properties, rainfall events and other weather variables and biological interactions in tropical humid ecosystems. Although, reliable estimates of crop water use can be obtained from local water balance studies, there is however, scanty data base on daily water requirement of field crops in tropics. This study focused on the identification of plant water potential and hydraulic conductance in relation to the soil-plant-atmosphere continuum (SPAC). Improved understanding of the basic biology of water ecophysiology of crops from tropical agroecologies will facilitate guided application of mineral nutrients to improve water use efficiency and overall crop yield. MATERIALS AND METHODS The whole plant hydraulic architecture (resistance in the flow pathway of water in the SPAC from the soil-root-stem-leaf), evaporative flux and plant water potential were examined in some cultivars of tomato and sweet melon grown in the screenhouse. The hydraulic conducganceplant water potential k (ψ) functions, (an index of transpiration induced water potential changes in the soil-leaf continuum), over the entire flow path was established towards the evaluation of hydraulic limits in the soil-leaf continuum. The hydraulic characteristics of melon and tomato was estimated from measurements of transpirational water loss rate (EL) and water potential (ψL) measured between pre- dawn and afternoon hours. During the sampling periods, transpiration flux and leaf water potentials were monitored. The soil and total plant resitance to water flow was calculated from slope of the relationship between water potential drop (change ψ ) and root water uptake (Q). Plant resistance was estimated for the measurements of transpirational water loss rate (EL) and water potential (ψL) day using the Ohm’s law analogy as Rp = (ψR -ψL)/Q. Transients and quasi-steady state mode were used in the measurement of conductance parameters (root, shoot, stem and leaf conductances), using a high pressure flow meter (HPFM, Dynamax, Houston). The theory of operation of the HPFM is discussed in more detail in Tyree et al. (1995). Through a watertight seal between the HPFM and the apical end of the plant selection, the HPFM was attached to the root and shoot system separately after cutting and clean shaven under water. A water-tight seal was produced using compression fittings manufactured by Dynamx. Water pressure was increased steadily from 0 to 0.5 Mpa at a constant rate of kPas-1, while measuring flow, F and applied pressure Pi every few seconds. For each parameter (root, shoot, stem and leaf), the slope of the relationship between F and Pi was taken as the hydraulic conductance (Tyree et al., 1995). The resistance element (R) was calculated from the inverse of K. Shoots were cut off while immersed in water and connected immediately to HPFM. The pressure was increased to 0.03 MPa and the shoot hydraulic conductance (Ksh) was measured in the steadystate mode (i.e. maintaining constant the pressure applied and measuring the corresponding flow). After about 30 minutes, leaves appeared to be infiltrated (perfused) with water as indicated by the change in colour and water dripping from the stomata), and the flow became stable. The hydraulic conductance of the leafy shoot was calculated as the ratio of the recorded flow to the pressure differences applied (P). The leaf blades were removed and the new hydraulic conductances of the leafless shoot (Kstem) was measured. Leaves can be regarded as having conductances in series with the stem because resistances (inverse of conductances) are additive in series, leaf hydraulic conductance (KI) was calculated by the difference in resistances as measured in leafy and leafless shoots (Tsuda and Turee, 2000). Total leaf surface area of each of the shoot was measured using a Delta T leaf meter. The cross-section of root and stem segments were measured using an electronic caliper. The hydraulic conductance parameters were scaled to root surface cross section area and leaf area of the shoot system. This was to enable examination of the physiological and ecological functions of the hydraulic conductances in terms of carbon cost to provide efficient hydraulic pathways and, hence, the adaptive advantage of a species/cultivar. The carbon efficiency of roots measurable in terms of root surface (cross section) area yields some measure of the cost of resource allocation (i.e., carbon investment by enhanced scavenging for water and mineral nutrients). RESULTS AND DISCUSSION Repeated measurement analysis of the parameters used to characterise plant water relations (1wp, EF and Kh) highlighted marked differences in melon and tomato. Figures 1 -3 show the day time courses (at 10, 12 and 1400 hours), while time component of predawn hour was included leaf water potential measurement) of leaf water potential, evaporative or transpiration fluxed (EF) and whole plant hydraulic conductance (Kh) obtained by EF method. Although significant species effects were obtained in the values of Iwp, EF and Kh, these parameters were sensitive to time and weather events of the day. Leaf water potential (Iwp) was charactersised by a decreasing trend from predawn values towards the afternoon hours in both melon and tomato (Fig. 1). Although melon displayed a more steady decrease compared to the sharp decline in tomato’s Iwp during the afternoon, lower values of leaf water potential were obtained at noon compared to 1300 and 1400 hours in both species. No important differences were obtained between tomato and melon in magnitudes of predawn leaf water potential. High predawn Iwp is attributable to tugor recovery from night time rehydration of the plant. The shapers of day time courses of EF and Kh were similar (Figs. 2 and 3), however, there were differences in their magnitudes from morning to mid- afternoon. The day time dynamics of EF and Kh showed increases in magnitudes from 1000 to 1200 h when these attributes attained peak values. Evaporative flux displayed steady increases in its magnitudes from early morning (1000 to 1400 h) (Fig. 2), while Kh declined gradually from sun rise to noon (Fig. 3). Daytime oscillation of water relation parameters may be sensitive to weather events of the day. Tomato had larger day time fluctuations in Iwp than melon, but the two species exhibited a similar trend in Iwp at midday (the characteristic midday depression of Iwp). The rate of change of Iwp during given interval of time during the day appear to relate to changes in EF and Kh. Daytime dynamics of Iwp, therefore, characterise the effects of species on water relations, as the effects of IWP was pronounced on daily cycles of hydration and dehydration. High predawn Iwp and low EF and K in the mornings when plants are close to full hydration, reverse trends in the magnitudes of these parameters in the afternoon hours (11-1400 h) following midday depressions of Iwp. Air temperatures and evaporative demand drive the diurnal course of Iwp and EF, the day time rate of change of EF relates to Kh and may be driven by weather events. Differences in daytime rate of changes of EF relates to Kh and may be driven by weather events. These daytime trends of Iwp between the species may also indicate differences in water consumption. Large canopy and vigour of growth induces greater plant water usage and positively influences EF and K in tomato. Tomato had higher IwP at predawn than at midday; the effects of high predawn Iwp positively translate to a higher midday Iwp in tomato. This may be explained by the differences in water absorption capacity of the root system compared to EF of canopy or a high Kh of the root system. Early in the morning, when temperature was mild, Kh did not keep pace with EF presumably due to high Iwp; towards midday period (Figs. 4 and 5), evaporative flux (EF) lagged behind hydraulic conductance but the deviation became pronounced from 1000 h to 1400 h. Superior within plant xylem transport and evaporative losses were obtained in tomato; from the relations of EF and Kh, this crop showed a higher hydraulic sufficiency than melon. The implications of hydraulic sufficiency for water use and productivity (carbon assimilation) in plants is reported in literature (Mencuccini, 2003; Stiller et al., 2003; Tyree, 2003). Physiological processes can explain the deviation of hydraulic conductance from transpirational water loss. Some amount of water is necessary to refill the dried tissues before onset of water loss via transpiration. The time lag between xylem water transport (sap flow) and transpiration would help to regenerate an increase in plant water content providing water for transport across the xylem conduits to rehydrate plant organs (Alarcon et al., 2000; Meinzer et al., 2003). The refilling of the water capacities of the tissues enhances the restoration of tissue turgor pressure and, hence, predawn leaf water potential. The contribution of water stored in plant tissues to the transpiration stream has been established (Nobel and Jordan, 1983; Meinzer et al., 2003). It is known that plant’s hydraulic transport efficiency regulates the diurnal and seasonal time scale of water loss; and leaf water status and such regulation is important to carbon assimilation, growth and survival under varying environmental growing conditions (Lo Gullo et al., 2003; Tyree, 2003). Since xylem characteristics are important to water transport in plants, a crop’s hydraulic architecture reveals the limitations placed by the resistance in the water transport pathway from the soil to root and leaf surface and, hence, plant productivity. Differences in water use among species/cultivars and the regulations of transpiration in response to soil moisture/climatic regimes can, therefore, be predicted through the theory of hydraulic limitations (Sperry et al., 2002). Soil and xylem account for a huge proportion of total path length from soil water source to evaporating surface in the leaf, therefore, the water potential of soil and xylem characteristics of the root and shoot system and, hence, the hydraulic conductance will exert influence on leaf water supply. The ease of water fluxes from the soil to the leaf (canopy) drives the architecture and physiology of plants. Whole plant water balance is, therefore, closely related to changes in component (root/stem/leaf) resistances (Nardini and Salleo, 2000; Tsuda and Tyree, 2000), with high hydraulic resistance from soil to the roots and the leaf blade and low resistance pathway in the stem. Although the resistance to water flow may be constant in a species, large ranges of behaviour occur between species during growth (Ruggiero et al., 2003). The ratio of leaf conductance (Kl) to total plant conductance (Kp) was remarkably high compared to other ratios (Figs. 6 and 7). In melon, it was over two-fold compared to the rations of Ksh, Kstem and Kr. In tomato, the ratio of Kl to Kp was over three-fold over the ratios of Ksh, Kstem and Kr to Kp. In addition to this, the values of Kr were compared to those Ksh and Kl; this gave higher ratio of Kr to Ksh compared to Kr and Kl in melon and tomato. The ratio of Kr to Ksh is two-fold in melon, it is over ten-fold in tomato, and in both species, the ratio of Kr to Kl is low. The greatest conductance was found in the leaf and the lowest in the root in both melon and tomato. While the highest within plant resistance to the flow of water is contained in the root system, the stem component constitutes the least resistance to within plant flow of water in tomato. The mechanisms underlying differences in water use among species may reside in changes in the magnitudes of component resistances in the water transport pathways of the soil-plant atmosphere interface (SPAC). Hydraulic characteristics may limit leaf water use efficiency in melon and tomato. Plasticity in hydraulic properties in a crop species may be offered through increasing resistance to water-stress induced xylem cavitations, this is central to the maintenance of an adequate water supply to the foliage (Melcher et al., 2003; Sobrado 2003). Tsuda and Tyree (2000) reported that the magnitudes of the resistance in the pathway of water transport is linked to the ability of a plant to maintain a favourable water status, and it may also imply improved efficiency of water use (Jones et al., 1982; Ruggiero et al., 2003). Parameters describing the hydraulic efficiency (architecture) of vascular system of tomato and melon were normalised in relation to plant attributes (Tables 1, 2, 3). The expression of hydraulic conductance of the root and shoot system relative to plant attributes (areas of leaf and stem cross sections) did not eliminate differences in the magnitude of conductance elements in tomato and melon. Differences between melon and tomato in whole plant leaf and stem area specific hydraulic conductance were obtained in this study. The description of hydraulic efficiency of in relation to plant attributes (areas of root surface and leaves; Kr/ rA, Kr/IA), yield some measure of the cost of resource allocation. There were variations in tomato and melon in terms of carbon investment for enhanced scavenging for water and mineral nutrients. Specific conductance parameters could relate to the necessities for modification of leaf area during growth to maintain constancy of hydraulic sufficiency to meet the water use demand by the canopy. The physiological and ecological functions of the hydraulic conductance in terms of carbon cost to provide efficient hydraulic pathways in the plant’s organs, could confer some adaptive advantage to a species. In plants, morphological or physiological adjustments are known to be associated with the maintenance of homeostasis of physiological functions (Gyenge et al., 2003; Sobrado, 2003; Tyree, 2003). Hydraulic architecture governs the pattern of canopy water use (transpiration rates), therefore, the sufficiency of the within plant xylem transport would have implications for efficiency of water use and productivity in the two species studied. Specific conductance parameters offer explanation for the necessities for modification of leaf area to maintain constancy of hydraulic sufficiency during growth in order to meet the water use demand of the canopy. In melon and tomato, crop water use and biomass accumulation (leaf area) adjust to water transport capacity, and higher conductance and higher stem/leaf area ratio (Huber values), were characteristics of tomato (size dependent functional modifications). High transpiration water loss requires high water uptake rates and, hence, the need for high hydraulic conductance to meet the demand of the canopy. Differences in the transport of water to leaf surfaces and separate control of water use, explains the tradeoff between water transport and leaf water use efficiency; an ecophysiological attribute with impact on the performance of a species in contrasting habitats (Gyenge et al., 2003; Sobrado, 2003). Studies have shown that leaf conductance positively relate to stomatal conductance, the stomata respond to Kl because changes in xylem conductance influences plant water status particularly leaf water status at the downstream end of the flow pathway (Three and Ewers, 1991; Hubbard et al 2001; Sobrado, 2003). The stomata respond to a variety of water relation parameters which are governed by a plant’s hydraulic architecture; therefore, gas exchange (stomatal conductance of gases) and, hence, carbon assimilation are responsive to changes in the hydraulic conductance of the soil to leaf pathway under varying environmental growing conditions (Hubbard et al., 2001; Stiller et al., 2003). The relationship between calculated whole plant hyradulic conductance (Kp measured with the evaporative flux method) and shoot conductance (Ksh measured on the HPFM) appears close. This is attributable to the fact that Kp includes the hydraulic conductance of the root system, which offers the highest resistance to water flow in a plant, and the frictional resistance of the proximal part of the crown. In addition, in both evaporative flux and HPFM methods, the hydraulic conductance elements were of higher magnitudes in tomato than melon. Plant hydraulic architecture is a function of xylem conduit dimensions (Tyree and Zimmermann, 2002; Cohen et al., 2003) and of the soil-root stele radial water pathway (Steudle and Heydt, 1997; Rieger and Litvin, 1999). These features which are genetically determined (Kavanagh et al., 1999), influence maximum leaf conductance compatible with maintenance of the sufficiency of the xylem system (Nardinin and Salleo, 2000). A high hydraulic conductance is known to be advantageous to plants in terms of efficiency of water and nutrient transport to leaves and, hence, high growth potential (as in the case with tomato). However, high hydraulic conductance due to wide and long xylem conduits might increase the vulnerability to caviation of plants (Tyree et al., 1994a; Nardinini and Tyree, 1999), and hence, the risk of xylem failure or dysfunction. Although, vulnerability to cavitation seems to place a limit on the minimum water potential than can be reached in the xylem without including catastrophic xylem dysfunction (Tyree and Sperry, 1989; Salleo et al., 2000), high specific leaf conductivities may facilitate the avoidance of xylem cavitation by prevention of xylem tension (pwp) from reaching levels that will cause cavitation/embolism. However, little is known of how vulnerable the xylem of most herbaceous species is to cavitation. On the other hand, if refilling or embolized xylem exists in these species possibly through mechanisms such as root pressure, recovery embolized xylem’s with therefore be important to recovery of productivity after episode of drought (Stiller et al., 2003). CONCLUSION Coupling of the root to shoot system (root-shoot communications), and whole plant regulation of water flux are present in melon and tomato. Under high transpiration/evaporative flux, total plant resistance is dominated by roots and leaf (canopy) resistance; a complex regulation that is difficult to dissect. The resistance of the vascular system influences water movement along gradient of total water potential, hence, high hydraulic conductance in tomato means that a relatively large gradient of water potential is needed to move enough water through the plant to maintain a given transpiration rate. The measurement of plant hydraulic parameters on whole shoot is referenced to their effect on water supply to leaves, this provides a comprehensive impressions of the actual hydraulic conductivity of the plants is obtained at maximum transpiration from the relation maximum day time evaporative flux and maximum diurnal drop in water potential (øpd-ø min). Some balancing between water transport capacity and evaporative flux to the supporting leaf area is evident. The sensitivity of leaf water potential offers osmotic regulation or protection, which could aim at maintaining constant hydraulic sufficiency in the species. The relation of water transport characteristics to plant variables/growth attributes indicates convergence of physiological functions in melon and tomato. REFERENCES
©2007, African Crop Science Society The following images related to this document are available:Photo images[cs07014t3.jpg] [cs07014t2.jpg] [cs07014t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}