 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
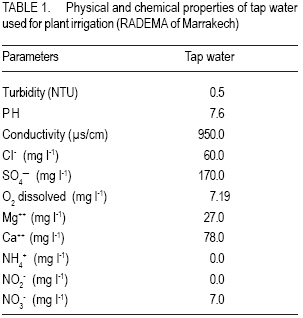
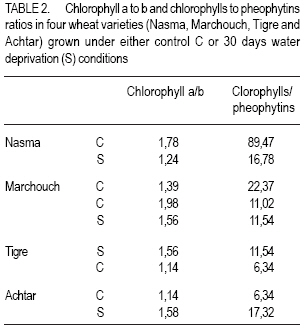
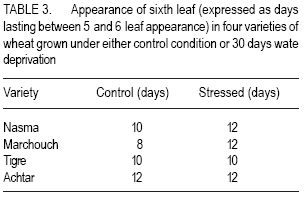
African Crop Science Journal, Vol. 15, No. 3, Sept, 2007, pp. 139-147 Water Deprivation Effect on Pigments and Proline Content, and Growth in Cultivated Wheat Varieties in Morocco Mimoun El Kaoau, Driss Hsissou And Abderrhmane Belhadri Université Cadi Ayyad, Faculté des Sciences et Techniques Received 1 August, 2004; accepted 29 December, 2006 Code Number: cs07015 ABSTRACT Plant growth and productivity are mostly affected by water shortage. This stress conditions induces plant cell dehydration, which in turn triggers varied adaptive mechanisms, manifested by molecular, biochemical and physiological changes. Water deprivation effects were evaluated in four water wheat (Triticum aestivum) varieties adapted to either irrigated (Nasma and Marchouch) or semi-arid (Tigre and Achtar) Moroccan regions. Pigments content (chlorophyll a and b, and pheophytins), membrane stability (assessed by electrolytes leakage), proline accumulation, and plant growth (assessed by sixth leaf appearance and third tiller development) were measured. In Nasma, chlorophyll b and pheophytin contents was observed. Moderate changes of these pigments contents were recorded for Marchouch and Tigre. In all varieties, water deficit caused an increase of electrolytes leakage and proline accumulation. However, these effects were more dramatic in the irrigationadapted varieties. Also, sixth leaf appearance was delayed and third tiller development was strongly inhibited in these varieties; whereas these parameters were almost unchanged in semi-arid environment adapted ones. Third tiller development is a good morphological criterion to assess water stress tolerance of the local wheat varieties studied. Key Words: Electroyle leakage, Triticum aestivum, water deficit RÉSUMÉ La croissance et la productivité de la plante sont surtout affectées par le manque d’eau. Ces conditions de tension entraînent la déshydratation des cellules de la plante, qui les détentes de virage change les mécanismes adaptifs, manifestés par les changements moléculaires, biochimiques et physiologiques. Les effets de privation d’eau ont été évalués dans quatre variétés de blé (Triticum aestivum) adaptées irrigation (Nasma et Marchouch) ou aux conditions semi arid (Tigre et Achtar) des régions marocaines. Le contenu de pigments (la chlorophylle a et b, et pheophytins), la stabilité des membranes (imposé par la fuite d’électrolytes), l’accumulation de proline, et la croissance de la plante (imposé par la sixièmes apparence de feuille et le tiers jusqu’à ce que le développement) ont été mesuré. Dans Nasma, le chlorophylle b et les contenus de pheophytin ont été observés. Les changements modérés de ces contenus de pigments ont été enregistrés pour Marchouch et Tigre. Dans toutes les variétés, tout déficit d’eau a causé une augmentation de fuite d’électrolytes et d’accumulation de proline. Cependant, ces effets étaient plus dramatiques dans les variétés d’irrigation adaptée. Aussi, la sixième apparence de feuille a été retardée et le tiers jusqu’à ce que le développement ait été fortement stoppé dans ces variétés ; tandis que ces paramètres étaient presque inchangés dans l’environnement semi aride. Le développement du troisième tiller est un bon critère morphologique pour évaluer la tolérance de tension d’eau des variétés de blé locales étudiées. Mots Clés: La fuite d’electrolyte, Triticum aestivum, déficit d’eau INTRODUCTION Plant growth and productivity are mostly affected by water shortage (Boyer, 1982; Kramer; 1983). This stress conditions induces plant cell dehydration, which in turn triggers varied adaptive mechanisms, manifested by molecular, biochemical and physiological changes (Cheeseman, 1988; Bohnert et al., 1995; Bray, 1997). Hence, cell elongation and division, tissue expansion (Gramier and Tardieu, 1999), leaf development (Sakuari et al., 1987; Abertnethy and McManus, 1998), and tiller formation (Davidson and Chevalier, 1987) can all be altered by water deficit. Some of these changed parameters such as membrane damage are used as criteria to evaluate drought tolerance (Premachandra and Shimada, 1988; Zuily-Fodil et al., 1990). The value of other parameters, such as the well studied proline accumulation, pigments content changes, and vegetative cycle variations, as criteria to estimate drought tolerance is not clearly established yet. Although the tolerance of higher plants to environmental stress is a complex multifactorial trait, it is still tempting to identify parameters that could accurately reflect this tolerance and facilitate the selection of tolerant varieties, especially in regions more prone to harsh environmental conditions. The objective of this study was to identify parameters of drought tolerance that could differentiate between two groups of wheat varieties possessing different water requirements. MATERIALS AND METHODS Plant material. Seeds of four bread wheat (Triticum aestivum L.) varieties that are adapted to irrigated conditions (Nasma and Marchouch) or semi-arid conditions (Tigre and Achtar) were provided by National Company for Seeds Marketing in Morocco. Seeds were initially germinated on moist filter paper placed in glass Petri dishes (9 cm in diameter) in the dark at 25 ± 1o C. Seedlings were transferred at the 1-2 stage on Zadokss scale (Zadoks et al., 1974) in plastic pots (24 cm in height and 20 cm in diameter) containing a mixture of peat, garden soil, and sand (2:1:1 v/v/v) with water saturation capacity of 75%. Four seedlings per pot and 15 pots for each variety and per condition were used. The pots were placed under natural day and night temperatures (20 ± 4 and 7 ± 3 oC, respectively) and under sunlight (10 ± 2h). All plants were irrigated every two days with 250 ml of tap water (properties on Table 1). After appearance of the fifth leaf, the pots were divided in two groups: control plants (continuous irrigation) and plants subjected to 30 days water deprivation (water saturation capacity drops to 5%). Water deprivation was imposed after the fifth leaf appearance since third tillers develop after this stage. For all subsequent experiments, material was gathered from leaves that develop during the stress period. Chlorophyll assay. After pigments extraction from 0.5 g of leaf fragments (collected from 20 plants from at least 3 pots) with 15 ml of acetone/ distilled water (90:10 v/v), the chlorophyll and pheophytin contents were determined spectrophtometrically (GBC UV/VIS 916) using a kinetic method of controlled pheophytinization (Laval-Martin, 1985). Addition of 5 µl of HCL (3 N) to 3 ml of the pigments was sufficient to produce total pheophytinization of both chlorophyll’s a and b. The pigments content was determined in three independent aliquots. Plant water content. Water content was determined for leaves collected from control plants (Qc) and water deprived ones (Qs) gravimetrically (drying done for 48 hr at 80oC). Electrolytes leakage. To measure electrolytes leagage, 0.2 g of leaves were washed with deionized water, cut in 0.5 cm fragements and put in test tubes contaning 40 ml of deionized water. Leaf fragments were shacked on a rotary shaker and washed three times with deionized water to remove the solution from the damaged cells (Blum and Ebercon, 1981). The leaf fragments were then placed in test tubes containing 15 ml of deionized water and shaken during 30 min. The conductivity (Lt) of the solution was then measured using a conductiometer (Orion 140). The maximum specific conductivity (Lk) of each sample was measured after shaking leaf fragments autoclaved at 120 oC during 30 min. The amount of electrolytes leakage of a stressed sample was given by 100 Lt/Lk. Measures were done on leaves from five different plants. Proline assay. Proline was determined five times in 0.2 g of leaf fragments collected from five different control well-watered plants or water deprived ones. Proline was assayed according to the method of Singh et al. (1973) modified by Paquin and Lechasseur (1979). The assay is based on the quantification of the reaction proline-ninhydrin by spectrophoteometric measurement. Proline reacts with ninhydrin by giving a colored complex. The intensity of the colour is proportional to the quantity of proline. Sixth leaf appearance and third tiller development. The date of appearance of sixth leaves (at its earliest stage) and third tillers (at 23 stage on Zadoks scale) were noted during development of 20 plants for each of the four varieties, under control or water deprivation conditions. The results were expressed in days lasting between fifth and sixth leaf, or as percentage of plants exhibiting third tiller. Statistical analysis. For the statistical analysis of data, two-way ANOVA (analysis of variance) was performed followed by the Turkey multiple comparisons test to determine significant differences among data. RESULTS Water deprivation effects on pigments content. Before treatment administration, there was no significant difference in chlorophyll a and b content between the four varieties (Fig. 1). After water deprivation, Nasma variety showed a larger decrease in chlorophyll a and b content. In fact, these pigments were reduced by half from 3.5 to 7.64 and from 2.8 to 4.28 µg g-1 dry mass (D.M.) for chlorophyll and b, respectively. Chlorophyll a and b slightly decreased in Marchouch and Tigre. In Achtar, chlorophyll a content was unchanged but chlorophyll b significantly decreased (Fig. 1). These changes are also illustrated by the ratio of chlorophyll a to b (Table 2). Indeed, after water deprivation, this ration significantly decreased in Nasma and increased in Achtar. On the other hand, this ratio was higher in semi-arid environment-adapted varieties Achtar (1.58) and Tigre (1.55), comparing to irrigated varieties Marchouch (1.36) and Nasma (1.24). Before water deprivation, pheophytin content was highest in semi-arid environment-adapted varieties Achar (1.66 µg g-1 D.M.) and Tigre (0.98 µg g-1 D.M.) compared with irrigated varieties Marchouch (0.53 µg g-1 D.M.) and Nasma (0.13 µg g-1 D.M.). After water deprivation, pheophytin content increased in Nasma (0.13 µg g-1 D.M.) but significantly decreased in Achtasr (0.49 µg g-1 D.M.). This pigment slightly decreased in Tigre and was unchanged in Marchouch (Fig. 1). In addition, after water deprivation, the ratio of chlorophylls to pheophytins decreased significantly and seems to stabilize around the same value in irrigation-adapted varieties Nasma and Marchouch (16.63 from 91.7 and 16.33 from 22.51, respectively (Table 2)). This ratio was lower for semi-arid environment adapted varieties and was unchanged by water deprivation in Tigre (11.6 from 10.98) but increased in Achtar (17.08 from 6.35). Plant water loss, electrolytes leakage and proline accumulation. Water deprivation induced a large decrease of plant water content in Nasma (36% water loss) and Marchouch (26% water loss). The semi-arid environment-adapted varieties, Tigre and Achtar, presented only 19.4% and of water loss, respectively (Fig. 2). Concomitantly, electrolytes leakage was more pronounced in Nasma and Marchouch than in Tigre and Achtar (Fig. 3). The difference between the two groups was observed since the first week of water deprivation and was maintained during the remaining stress period. Also, after 30 days of water deprivation, proline accumulation increased greatly in leaves of all four varieties. However, the irrigationadapted varieties, Nasma and Marchouch, showed twice as higher a proline accumulation (46.73 and 47.32 μg g-1 D.M., respectively) compared with semi-arid environment-adapted varieties Tiger and Achtar (20.21 and 26.5 μg g-1 D.M., respectively) (Fig. 4). Sixth leaf appearance and third tiller development. Water deprivation delayed appearance of sixth leaf by two days in Nasma and four days in Marchouch varieties. In Tigre and Achtar varieties, sixth leaves appeared at the same time as in control conditions (Table 3). Third tiller development was strongly inhibited by water deprivation in irrigationadapted varieties, Nasma and Marchouch. Indeed, only halfway through water deprivation period (about two weeks), less than 30% of plants from these varieties developed their third tillers (Fig. 5). This inhibition was maintained during the entire water deprivation period. Contrary to this observation, semi-arid environment adapted varieties, Tiger and Achtar, continued to develop third tillers even after a prolonged water deprivation treatment. Effectively, after 24 days of water stress, at least 80% of plants presented third development (Fig. 5). DISCUSSION Water deprivation-induced stress had different quantitative and qualitative effects on the parameters measured in four wheat varieties cultivated in Morocco, adapted to whether irrigated or semi-arid environments. The changes of some parameters were extreme in Nasma and Achtar varieties, while intermediate effects were recorded for Marchouch and Tigre. For instance, chlorophyll a and b contents, decreased significantly in Nasama and moderately in Marchouch and Tigre. In Achtar, the chlorophyll b decreased significantly while chlorophyll a was unchanged. In addition, after water deprivation, the ratio of chlorophyll to pheophytins decreased significantly in irrigation-adapted varieties and increased, especially in semi-arid environmentadapted Achtar. Since pheophytins are represented by very low quantities, this indicates that overall, the chlorophyll pigments content is decreased by water deprivation. Previous studies have shown droughtinduced decrease in pigments content in many plants including pea (Moran et al., 1994), Nerium oleander (Demming-Adams et al., 1988), and durum wheat (Loggini et al., 1999). The decrease of chlorophyll a and b contents recorded in Nasma, Marchouch, and Tigre confirms these findings. Besides, it is suggested that drought-tolerant varieties might have greater capacity to preserve their photosynthetic apparatus (Anderson and Aro, 1994; Loggini et al., 1999). In semi-arid variety Achtar, the content of chlorophyll a, the principal constituent of PSI and PSII reaction centers, was unaffected by water deprivation and hence seems to support this hypothesis. Proline accumulation has also been well documented in osmo-stressed plants, and different hypotheses have been proposed to explain this fact. In effect, proline is suggested to act as an organic solute, directly participating in osmotic adjustment after osmotic stress, and/or protect macromolecules after environmental perturbations (reviewed by Delauney and Verma, 1993). The high proline increase observed in our irrigation-dependent varieties Nasma and Marchouch can be correlated with a higher water loss and high electrolytes leakage in these varieties compared with their semi-arid environment-adapted counterparts. This rather suggests that proline accumulation is tributary of membrane integrity and acts as a signal to downstream events involved in osmoadjustments, but this remains to be confirmed. However, very recent findings (Huguet-Robert et al., 2003) showed that osmo-induced proline accumulation is inhibited by polyunsaturated fatty acids (important constituent of cell membrane) and, therefore, further supports dependence of proline accumulation on cell membrane stability. Most important, water deprivation had a marked inhibition effect on third tiller development in irrigation-adapted varieties, Nasma and Marchouch. This inhibition was so effective that after only 15 days of water deprivation, 70% of plants failed to develop their third tillers. Interestingly, third tiller development was almost unaffected in semi-arid environment-adapted Tigre and Achtar. Therefore, the third tiller development parameter can be considered as a morphological criterion to differentiate between the two types two types of varieties studied. In addition, Ceccarelliu and Grando (1991) suggested that drought tolerance might be correlated with higher productivity in barley, whereas Kirkham and Kanemasu (1983) claimed that there is a negative relationship between tolerance and the yield potential. Our results clearly demonstrate that the semi-arid local varieties (Tigre Achtar) tolerated better water deprivation and developed their third tillers. Therefore, these “water-deficit tolerant” varieties will definitely have a greater yield potential. CONCLUSION Water deprivation-induced pigments content changes are important but not sufficient to discriminate between different types of varieties. Also, proline accumulation is rather a better indicator of membrane stability than drought tolerance. Among the parameters studied, we can confidently consider that third tiller development is a better criterion to estimate water deprivation sensitivity for the wheat varieties used in this study. ACKNOWLEDGEMENTS Part of this work was supported by the Biology Department (Faculté des Sciences et Techniques, Marrakech) and PROTARS project no p5ti/03. We are grateful to professor G. Bitton (University of Florida) and Dr. A. Boussaid for reading the first draft of the manuscript. We also thank Director REFERENCES
©2007, African Crop Science Society The following images related to this document are available:Photo images[cs07015t1.jpg] [cs07015t2.jpg] [cs07015t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}