 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
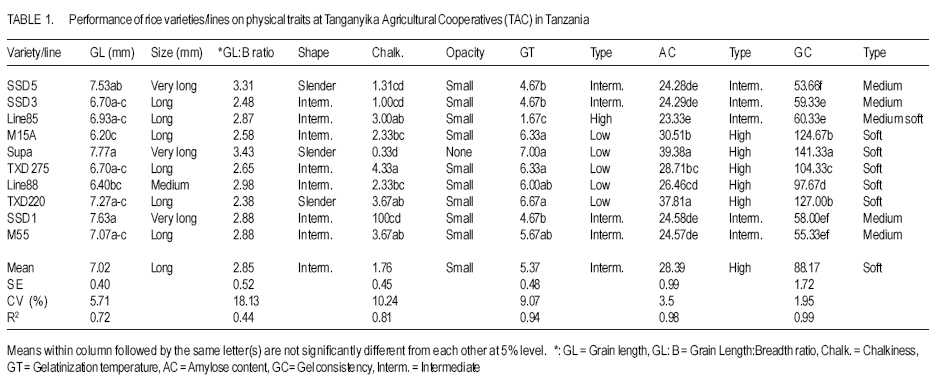
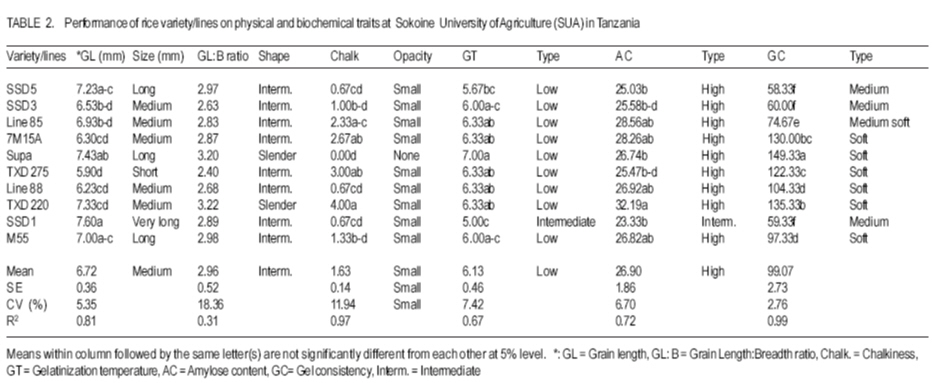
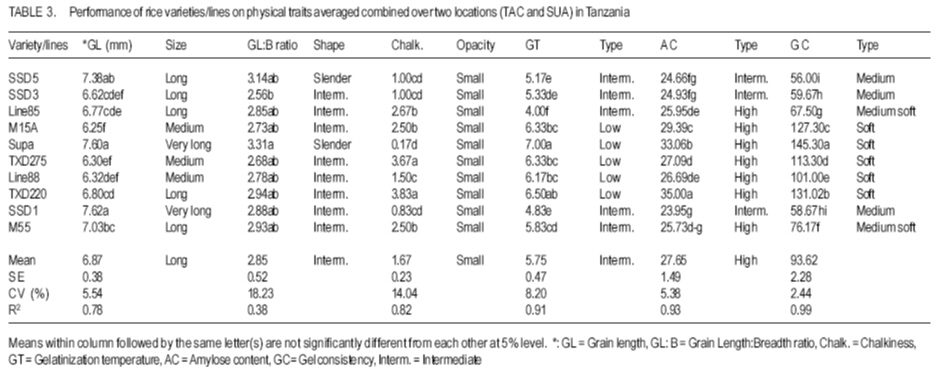
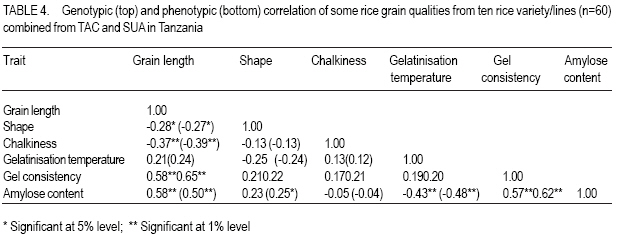
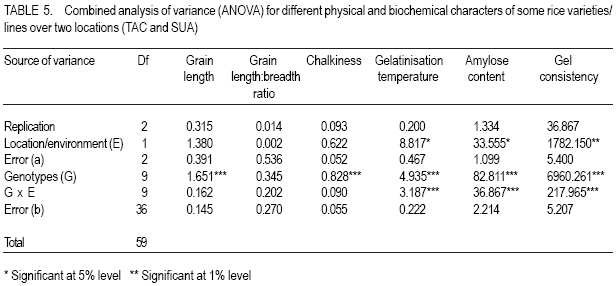
African Crop Science Journal, Vol. 15, No. 4, Dec, 2007, pp. 173-182 Influence of Genetic and Genotype X Environment Interaction on Quality of Rice Grain M. Nkori Kibanda and A. Luzi-Kihupi1 Agricultural Research Institute, KATRIN, P/Bag, Ifakara, Tanzania Code Number: cs07018 ABSTRACT Rice (Oryza sativa) grain attributes, including among others milling quality, grain length and shape, amylose content, and aroma are critical in varietal development and subsquent adoption at the farm level. It is, therefore, important to understand the influence of genetic and G x E interaction on these grain attibutes. Thus, experiments were laid out during 1999-2000 at the Tanganyika Agricultural Cooperative, Ifakara (irrigated culture) and at Sokoine University of Agriculture (upland conditions) both in Morogoro region, to obtain relative magnitudes of variety x environment interactions, heritability and genetic advance aspects on physical and biochemical rice grain quality attributes. High significant positive genotypic and phenotypic correlations were revealed between gel consistency (GC) and both the grain length (GL) and amylose content (AC). Gel consistency had high estimates of heritability and expected genetic advance, and is thus a reliable selection criterion for amylose content in early generations of rice improvement. The observed variation among genotypes under different environments suggest that in order to ensure high grain quality, there is need to select genotypes for particular cropping environments. Key Words: Genetic advance, heritability, Oryza sativa, upland, Tanzania RÉSUMÉ Les attributs de riz (Oryza sativa), inclut parmi d’autres la qualité au moulant, la longueur et la forme des grains, le contenu d’amylose, et l’arôme sont importants dans le développement variétal et l’adoption au niveau de ferme. C’est plus important de comprendre l’influence de génétique et l’interaction G x E sur ces attributs de grain. Ainsi, les expériences ont été faites entre 1999-2000 à la Coopérative agricole de Tanganyika, Ifakara (sous culture irrigué) et à l’Université d’Agriculture de Soko¿ne (sur haute terre conditionnée) les deux dans la région de Morogoro, pour obtenir des magnitudes relatives des interactions variété x environnement, héritabilité et les aspects d’avance génétiques sur les attributs de qualité de grain de riz physiques et biochimiques. Les hautes corrélations, significatives et positives entre génotypes et les phénotypes ont été révélées entre l’homogénéité de gel (GC) et la longueur de grain (GL) et le contenu d’amylose (AC). L’homogénéité de gel a eu la haute estimation de héritabilité et l’avance génétique prévue, et est un critère de sélection fiable pour le contenu d’amylose dans toutes les générations d’amélioration de riz. La variation observée parmi les génotypes sous les environnements différents suggère que pour garanti la haute qualité de grains, il y a nécessité de choisir des génotypes particulier pour des environnements différents. Mots Clés: Avance génétique, héritabilité, Oryza sativa, haute terre, Tanzanie INTRODUCTION Rice (Oryza sativa) grain quality attributes are among important factors prior to variety adoption. They include physical (milling quality, grain length and shape) and biochemical (amylose content, gel consistence, gelatinisation temperature and aroma) characteristics. Such grain quality attributes vary among varieties and production environments (Unnevehr et al., 1992; Jennings et al., 1979; Fan et al., 1999). Post harvest factors like rice parboiling, aging or storage for 3-4 months were reported to improve total and head rice (Juliano, 1972). In accordance with the classification by IRRI (1988), extra long (6.61-7.50 mm) to medium (5.51-6.60 mm) grain lengths are preferred in Tanzania. Correlation studies among physical and biochemical characteristics show that grain length have negative associations with grain width and shape (Juliano and Villareal, 1993) and chalkiness (Khush, 1994). Inheritance studies further indicate that grain length is highly heritable, and its inheritance varies from monogenic to essentially polygenic (Mackill et al., 1996). The level of chalkiness affects milling quality as it provides weaker points for breakage under mechanical milling. Low and high temperatures after flowering, and excess soil fertility in rainfed lowland are factors for increased chalkiness (Resurreccion, 1977). The trait is quantitatively inherited and its mode of inheritance depends on parental and environmental influences (Chang and Somrith, 1979). According to Chang and Somrith (1979) other breeders’ experiences show that chalkiness fixes earlier and therefore, strict selection for chalkiness should begin from the early generations. Amylose is a factor responsible for cooked rice hardness, stickiness, colour, gloss and general acceptability (Mackill et al., 1996). Low ambient temperatures during ripening increase amylose content and decreases with increased nitrogen application (Juliano, 1972; Resurreccion, 1977). Depending on amylose content, rice vary in water requirements and texture after cooking. In Tanzania, intermediate amylose content (21-25%) rice that becomes fluffy when cooked and remains soft when allowed to cool is generally preferred. Amylose content has been reported to show positive correlations with gel consistency and gelatinisation temperature, but a negative correlation with grain width (Juliano and Villareal, 1993). The inheritance of amylose content was reported to be simple (Mackill et al., 1996). Gelatinisation temperature (GT) was documented to vary with respect to the ratio of amylose to amylopectin, starch crystallinity, granule size, distribution and the amount of minor constituents like phosphorus, lipids, protein and enzymes (Chattakanonda et al., 2000). High air temperature after flowering raises GT; while low air temperature lowers it (Mackill et al., 1996). Rice with high GT tend to require more water and time to cook than those possessing either low or intermediate GT (Chatterjee and Maiti, 1985). Juliano Rice varieties possessing intermediate GT 70-74°C are preferred in the country. Juliano (1987) also revealed that high GT was associated with low amylose. The inheritance of gelatinisation temperature is, however, simple involving one or two major genes (Shan et al., 1987). Gel consistency depends on the variations in amylopectin fractions (Juliano and Perdon, 1975). Addition of nitrogenous fertiliser at heading stage somehow increases the protein content, which subsequently contributes to harder gel consistency (Seetanum and De Datta, 1973). Some high GT rice tends to give hard cooked rice and rice products due to the presence of large amylopectin molecules (Perez et al., 1979). In Tanzania, soft gel consistency (61-100 mm) is preferred to hard gel consistency. Effects of environmental factors on rice grain qualities tend to limit the effectiveness of selection of superior genotypes. Genotypes x environment interactions reveal the need for development of genotypes that should be tested and selected for specific growing environments (Mehta et al., 1984; Fehr, 1987). Tanzania has varied rice growing environments for which recommendations of varieties are being made. Recently, various rice breeding materials of different yield potentials and grain qualities have been developed. Apparently, there is no information on the effect of rice grain qualities due to genotypes x environments (ecosystems) interactions. However, experiments conducted elsewhere are insufficient and not representative of the available genotypes under the existing varied environmental conditions. The present study was therefore, aimed at evaluating the performance of the new rice genotypes and their interactions on grain quality attributes under varying environments (upland rice and lowland conditions). The specific focus was on assessing the contribution and the relative magnitudes of rice variety and variety x environment interaction on physical and biochemical traits under upland and irrigated ecosystems; and estimating and assessing correlations and genetic parameters of physical and biochemical grain quality attributes and determining their relative importance for rice improvement under upland and irrigated cultures MATERIALS AND METHODS The experiment was conducted during 1999-2000 at the Tanganyika Agricultural Cooperatives (TAC), Ifakara (irrigated), and Sokoine University ofAgriculture (SUA) Farm (upland) in Morogoro region. Ten rice varieties/lines including SSD1, SSD3, SSD5, M15A, Line85, Line88, TXD275, TXD220 and Supa (control), and some amutant from SUA were used in the study. Seeds tested were selected on the basis of their potential performance for future seed release. The study was laid out in a randomised complete block design (RCBD) with three replications and plot sizes of 4 m x 2 m. Seeds (2 - 3) were dibbled per hill at spacing of 20 cm x 20 cm. Thinning was done to two seedling/plant and the harvested area was 3.5 m x 1.5 m. Weeding, N fertiliser application and other agronomic practices were done as per recommended standards. Data recorded comprised grain length and width, grain shape and opacity, amylose content, gel consistency, gelatinisation temperature and aroma. A graphic (logarithmic) paper method described by Jennings et al. (1979) was used to determine grain size, shape and opacity of each genotype. Amylose content was determined as per modified simplified assay (manual) procedure by Juliano et al. (1981). A method as described by Cagampang et al. (1973) was used to determine gel consistency of rice genotypes. Opacity of milled rice was visually rated in terms of chalky proportions of the grains. The assessment was done on any chalkiness, whether white belly, white centre or white back. All grain quality attributes results were rated, evaluated and assessed according to the scale established by IRRI (1988). Statistical analyses were performed per site as well as when pooled over two locations. SAS Software, Version 8.12 (1997) was used for this purpose. Genotypic and phenotypic correlation coefficients were computed by a method proposed by Miller et al., 1958; Steel and Torrie, 1980. RESULTS AND DISCUSSION At TAC site which was under irrigated condition, soils were medium acidic clay, with medium Ca+, Mg+ and K+ and low Na+. Temperatures ranged from 31.5 -27.1°C (maximum) and 23.3 -19.6°C (minimum). At flowering, temperatures ranged from 29.2-28.3°C (maximum) and 23.1-21.7°C (minimum). A rainfall dependent SUA site had its soils as slightly acidic clay sand with medium Ca+, K+ and Na+ and high Mg+. Temperatures ranged from 30.6 -17.4°C. Mean rainfall during the entire growing season was about 140 mm month-1. Performance results of physical and biochemical traits of lines/variety tested are shown in Tables 1, 2, 3, and 4. Environmental means for grain length and amylose content were higher at TAC than at SUA site, but the reverse was true for gelatinisation temperature and gel consistency at SUA than at TAC site. The significant difference in grain size and non-significant performance in shape among genotypes at TAC and SUA sites suggest differences in inherent genetic characteristics and genotypic responses due to environmental changes over the trait within locations. Similar results on high and less variability of the respective grain size and shape under different environmental regimes have been reported (Unnevehr et al., 1992). Genetic variability has also been reported (Jennings et al., 1979). The significant difference that was depicted on grain shape, when data were pooled, suggests effect of a pronounced environmental influence of grain shape among rice genotypes. Grain size ranged from very long to medium with slender to intermediate shape at TAC, whereas grain size at SUA were long ranged from long to short with intermediate shape. In a combined analysis grain size ranged from long to medium with slender to intermediate shape. Although chalkiness was significant (P≤0.05), it was regarded as negligible in both the environments tested. There were significant differences (P≤0.05) in gelatinisation temperatures among genotypes in all the environments tested (Tables 1, 2, and 3). Except for Line 85 at TAC, which had high gelatinisation temperatures, the rest demonstrated intermediate to low GTs at SUA and when data were combined. Most entries recorded low gelatinisation temperature (GT) at SUA than at TAC, and this was due to temperature differences between locations. Results imply that variations in GT could be influenced by genetic differences and the genotypic differential responses within and between locations. Similar findings on genetic variability in rice have been documented (Fan et al., 1999), while influences of high or low temperatures above or below 30°C - 33°C during flowering have been documented to cause differential low and high gelatinisation temperature, respectively (Yoshida, 1981; Mackill et al., 1996). The GT values in all the locations ranged from 1.67 (low) only for Line 85 to 7.00 (low) for Supa at TAC and 5.00 (intermediate) to 7.00) at SUA. Similarly, GT were 4.00 for Line 85 to 7.00 for Supa when data were pooled. Significant (P≤0.05) differences on amylose content among tested genotypes were realized from TAC and SUA sites, even when data of two locations were pooled (Tables 1, 2, and 3). Amylose content among genotypes ranged from 23.33 - 39.38 for Line 85 and Supa varieties at TAC, 23.33 - 32.19 for SSD1 and TXD 220 at SUA sites all falling between high to intermediate types (Tables 1, 2, and 3). Variations in amylose content suggests the existence of genetic variabilities. Genetic effects on the performance of amylose content in rice have been reported elsewhere (Mackill et al., 1996), while amylose content increase due to increases in temperature up to 29°C, above which amylose content decline was documented (Resurreccion et al., 1977). Significant differences (P≤0.05) in gel consistency among tested genotypes were obtained from TAC and SUA sites and when data were pooled up (Tables 1 and 2). Gel consistency among varieties ranged from 53.66-141.33 at TAC and at SUA sites and from 56.00-145.30 when data were combined. From all the sites and in the combined analysis, maximum and minimum ranges were obtained from the respective SSD5 and Supa varieties. Higher values were noted at TAC than at SUA. Results from treatment means and the combined analyses of variance revealed that rice varieties, environments and variety x environment interaction were probable key attributes to the differential performance of gel consistency (Table 3). Differences in performance in gel consistency of rice varieties suggest the existence of genetic variability. Juliano and Perdon (1975) reported a similar finding on existence of genetic variability on the performance amylose content in rice genotypes. All genotypes tested possessed medium gel (41-60 mm) to soft gel consistency (>80 mm) and differences due to temperature and soil moisture regimes among rice varieties. Since rice with long to intermediate grain size, with slender to intermediate shapes that are translucent with low to intermediate gelatinisation temperature, high amylose content and soft gel consistency are preferred, results suggest that the rice breeding materials tested are likely to be adopted for cultivation in Tanzania (Tables 1, 2, and 3). From the combined analysis of variance (Table 5), genetic variance was higher than location and genotype x environment interaction variances for all the characters except for gelatinisation temperature. There were no environmental influences on grain length, grain size and chalkiness. Although environmental variance component was unimportant to grain shape, the genotype x environment was very important for gelatinisation temperature, amylose content, and gel consistency, suggesting that environmental factors were also responsible for the modification of amylose. Similar results on the influence of temperatures below/beyond an optimum 30 °C33°C were also reported to decrease or increase gelatinisation temperature (Mackill et al., 1996) and gel consistency. Inconsistent treatment results have demonstrated the importance of environment in the general performance of all the traits except for grain shape. The scenario signals difficulties in selection of all the traits evaluated. A more systematic approach aimed at separating environmental effect from genetic component is required to predict success in selection during crop improvement. Results of genotypic and phenotypic correlations of a combined analysis are given in Table 4. Amylose content was negatively and significantly (P<0.01) correlated with gelatinisation temperature, but positively and significantly (P<0.01) correlated with gel consistency. This implies that improving amylose content would simultaneously increase gel consistency, while discriminating gelatinisation temperature. Positive correlations between amylose content with gel consistency and negative correlations with gelatinisation temperature have been obtained from world rices (Juliano and Villareal, 1993). Results of variance components for grain quality characters of ten genotypes combined over two environments are shown in Table 5. The genetic variance was higher than location and genotype x environment interaction variances for all the characters, except for gelatinisation temperature. However, genotype x environment was very important for amylose content, gelatinisation temperature and gel consistency suggesting influence of environmental changes in performance of the three mentioned rice traits. From the variance components for grain quality attributes over two locations, all traits had high heritability except for grain shape. Amongst them, gel consistency was the only trait that was observed with both highest broad sense heritability and expected genetic advance. In addition, the trait exhibited significant positive genotypic and phenotypic correlation coefficients with amylose content, which also is the most important trait that greatly determines the eating quality of rice. CONCLUSION Environments appreciably affect the performance of rice genotypes and retard selection progress for grain quality improvement. Despite their inconsistent performance for grain quality traits over two contrasting environments, all tested genotypes possess a wide range of acceptable grain quality attributes and, therefore, stand a chance of being adopted by rice farmers and consumers in Tanzania. In terms of genetic values of the rice traits, rice breeders should consider gel consistency as a reliable indirect selection criterion for amylose content improvement in early generations of rice breeding program. ACKNOWLEDGEMENT We sincerely acknowledge the Ministry of Agriculture for funding this project. We also wish to thank technicians of the Agricultural Research Institute – KATRIN, Morogoro, Tanzania and those from Sokoine University of Agriculture (SUA), Morogoro, Tanzania for successful laboratory analyses of the grain quality attributes. REFERENCES
©2007, African Crop Science Society The following images related to this document are available:Photo images[cs07018t2.jpg] [cs07018t1.jpg] [cs07018t3.jpg] [cs07018t4.jpg] [cs07018t5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}