 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 15, No. 4, Dec, 2007, pp. 191-199 Etude de la Résistance au Flétrissement Bactérien de Lignées Inter-Et Intraspécifiques de Riz de Bas-Fonds en Conditions D’infestation Artificielles S. L. Ouedraogo, I. Somda1, I. Wonni1 et Y. Seré2 Institut de l’Environnement et de Recherches Agricoles (INERA) Station de Farakoba BP 910 Bobo
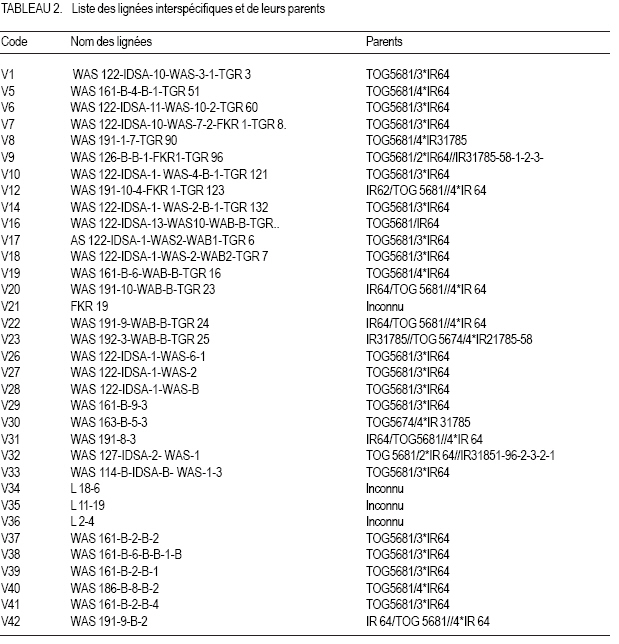
Dioulasso, Burkina Faso Code Number: cs07020 RÉSUMÉ Le riz (Oryza sativa) est une nourriture de base importante au Burkina Faso. Cependant, la productivité de riz au Burkina Faso est affectée par l’infestation bactérienne de flanche menant à l’échec total de récolte dans beaucoup des cas. Il y a donc urgence d’évaluer et par la suite introduire des cultivars résistants pour toutes les zones écologiques du Burkina faso. Dans cette étude, cette évaluation de la résistance à la flanche bactérienne a été faite en utilisant soixante- neuf lignées de riz des basses terres, dont huit lignées intra spécifiques et soixante et interspécifiques. Une variante de Xanthomonas oryzae pv. oryzae du Burkina Faso a été utilisé pour tester les lignées candidates. Une variante de référence (NCPPB 1150) a été obtenue au Danemark et de l’eau distillée a été utilisée respectivement comme le contrôle positif et négatif. La variété susceptible à la bactérie flanche, la variété chinoise TCS 10 et la résistante FKR19 ont été aussi utilisées comme contrôle. L’expérience a été conduite en serre à la température de 28-30°C et 12 heures de cycle de lumière. La conception expérimentale était du type split plot avec trois réplications. Apres quinze jours les vieilles plantes de riz ont été vaccinées et les symptômes de maladie ont été observés quatorze jours après l’inoculation. Les résultats ont indiqué que 55.07 % des lignées sont résistantes, 23.20 % modérément résistant, 5.80 % modérément susceptible, 7.24 % susceptible et 8.69 % extrêmement susceptible. Il a été aussi observé que 78.68 % de lignées interspécifiques étaient plus résistantes en comparaison avec 75 % d’intra spécifiques. La variante bactérienne de Bagre (Burkina Faso) était plus virulente que le NCPPB de référence 1150. Les lignées inter spécifiques et intra spécifiques doivent être essayé dans les conditions de champ avant la recommandation pour l’adoption dans les zones infestées par Xanthomonas oryzae pv. oryzae. Mots Clés: Bactérien flanche, Burkina Faso, des basses terres, la résistance, Oryza sativa ABSTRACT Rice (Oryza sativa) is an important staple food in Burkina Faso. However, rice productivity in Burkina Faso is affected by bacterial wilt disease leading to total crop failure in many instances. Therefore, there is need to evaluate and subsequently introduce cultivars that exhibit resistance under ecological zones in Burkina Faso. In this study, evaluation of the resistance to bacterial wilt was done using sixty nine lines of low-land rice, eight of which were intraspecific and sixty one interspecific. A strain of Xanthomonas oryzae pv. oryzae isolated from Burkina Faso was used to challenge the candidate strains. A reference strain (NCPPB 1150) obtained from Denmark and distilled water were used as positive and negative control, respectively. The susceptible variety to bacterial wilt, chinese variety TCS 10 and the resistant one, FKR19 were also used as control. The experiment was conducted in screenhouse at the temperature of 28-30 °C and 12 hours daylight cycle. Experimental design was a split splot with three replications. Fifteen days old plants of rice were inoculated and disease symptoms were observed fourteen days after inoculation. The results indicated that 55,07 % of the lines are resistant, 23,20 % moderately resistant, 5,80 % moderately susceptible, 7,24 % susceptible and 8,69 % highly susceptible. It was also observed that 78,68 % of interspecific lines were more resistant compared to 75 % of the intraspecific. Bacterial strain isolated from Bagre (Burkina Faso) was more virulent than the reference NCPPB 1150. The inter and intraspecific lines need to be tested in the field conditions before recommendation for adoption in zones infested by Xanthomonas oryzae pv. oryzae. Key Words: Bacterial wilt, Burkina Faso, low-land, resistance, Oryza sativa INTRODUCTION Le flétrissement bactérien du riz (Oryza sativa L.), causé par Xanthomonas oryzae pv. oryzae a été signalé aux Philippines depuis 1950 (Swings et al., 1992 cités par Mew et al., 1994). Cette maladie est devenue importante à partir des années 60 quand les variétés à haut rendement ont été adoptées. Elle est, de nos jours considérée comme l’une des maladies la plus dévastatrice du riz en Asie (Agarwal et al., 1994 ; Sy et Seré, 1996). D’après Robert et Pamela (1992), les pertes de rendement occasionnées par cette maladie sont de 20-30% et peuvent atteindre jusqu’à 50%. En Inde, ces pertes atteignent 74-81,3% selon les variétés et la saison de culture (Ahmed et Sing, 1975 ; Sing et Rao, 1977). Aux Philippines, avec la découverte de variétés résistantes à certaines souches de Xanthomonas oryzae pv. oryzae telles que la TKM-6, Sigadis et Cas 209, l’IRRI a procédé à l’incorporation de gènes de résistance à de nouvelles variétés de riz (Mew et al., 1992). En Afrique, notamment au Sahel, le flétrissement bactérien a fait son apparition pour la première fois au Mali en 1979 (Buddenhagen et al.,1979). Il a été ensuite signalé au Sénégal en 1980, au Burkina Faso en 1981 et au Niger en 1983 (Institut du Sahel, 1991) . Au Burkina Faso, l’agent pathogène a été identifié en 1982 (Seré et Nacro, 1992). Cependant, ces deux auteurs indiquent en (1996), que les pertes causées par la maladie sont peu évaluées en milieu paysan. Elle a été la maladie la plus dévastatrice du riz sur la plaine rizicole de Bagré au cours de ces dernières années où 50 % de dégâts ont été enregistrés en 1998 (Ouédraogo et al. 2004 ; Kaboré et Ouédraogo, 2004) . La variété TCS 10 reconnue pour sa bonne productivité, mais très sensible à la bactériose est de plus en plus abandonnée par les producteurs au profit de la FKR 19 et FKR 14. Malgré la présence de la maladie et les pertes qu’elle engendre, aucune méthode de lutte efficace et adoptable par les producteurs n’est disponible. Pour contribuer à la résolution de ce problème, la présente étude se propose d’évaluer la résistance au flétrissement bactérien de lignées inter- et intraspécifiques de riz de bas-fonds en conditions d’infestation artificielles. MATERIEL ET METHODES Matériel végétal. Il est composé de 08 lignées intraspécifiques (Tableau 1) et de 61 lignées interspécifiques (Tableau 2) de riz de bas-fonds provenant de l’ADRAO/ Sénégal, de l’ADRAO / Côte D’Ivoire et du CIAT/Colombie. Matériel biologique. La souche de référence Xanthomonas oryzae pv. oryzae NCPPB 1150 a été fournie par le laboratoire de bactériologie de l’Institut Danois de Pathologie des Semences. La souche XBFB (Xanthomonas oryzae pv. oryzae de Bagré, Burkina Faso) a été isolée à partir des plants de riz infectés de la variété TCS 10 collectés sur le périmètre rizicole irrigué de Bagré. Méthode de détection et d’isolement. Les feuilles de plants de riz flétris prélevées au champ et où l’on soupçonne la présence de la bactérie, ont été coupées en petits morceaux d’environ un cm et examinés au microscope entre lames et lamelles dans une goutte d’eau distillée stérile. En cas d’infection, on observe des bactéries qui exsudent des tissus végétaux. Les fragments infectés sont donc retirés du microscope et laissés pendant 10-15 mn dans un petit becher contenant quelques ml d’eau distillée stérile afin d’obtenir plus de cellules bactériennes dans l’eau. A l’aide d’une anse, la suspension est prélevée du becher et étalée sur le milieu de culture (Facteur de Croissance (FG). La souche de référence est aussi étalée sur le même milieu pour comparaison. Les boîtes de Pétri sont incubées en position inversée à la température de 25-30° C pendant 4872 heures. L’isolement se fait sous la hotte à flux laminaire. Purification et identification des colonies isolées. Les colonies suspectes de Xanthomonas oryzae pv. oryzae sont purifiées sur le milieu King’s B. La souche de référence NCPPB 1150 est aussi étalée sur le même milieu pour comparaison. Les tests biochimiques standards utilisés pour l’identification des bactéries après purification sont : a) Réaction de Gram (Test de solubilité dans KOH, Lelliot et Stead, 1987). A l’aide d’un cure dent stérile, les bactéries sont prélevées à partir d’une culture pure de 24-48 heures et frottées rapidement dans une goutte d’une solution aqueuse de 3% de KOH préalablement déposée sur une lame de verre. Au bout de quelques secondes, on soulève le cure dent de quelques cm au dessus de la lame. S’il se forme un filet visqueux, la bactérie est Gram négatif. Les bactéries Gram positif ne produisent pas de filet. b) Test d’oxydase (Kovac’s, 1956). Une solution aqueuse de 1% de dihydrochlorure tétramétylparaphénylènediamine a été préparée. Une goutte de cette solution est déposée sur du papier Whatman. Un prélèvement de culture bactérienne de 24-48 heures est effectué à l’aide d’une anse et frottée dans la goutte. Si le réactif vire au pourpre de façon instantanée ou dans les 10 secondes qui suivent l’application de la culture, la réaction est positive. Dans le cas contraire, elle est considérée négative. c) Oxydation/ Fermentation (O/F) (Fahy et Persley, 1983). Pour chaque isolat à tester, deux tubes contenant le milieu O/F sont inoculés à l’aide d’une culture bactérienne de 24-48 heures. L’inoculation consiste à introduire une anse pleine de bactéries directement au fond du tube à essai et à la retirer aussitôt. L’un des tubes est recouvert d’huile de paraffine d’un à deux cm d’épaisseur afin d’empêcher toute infiltration d’oxygène. La bactérie de référence est utilisée comme témoin positif. Les tubes sont mis en incubation à 28°C et les observations sont faites quotidiennement à partir du troisième jour jusqu’à deux semaines. Un changement de couleur de vert olive au jaune dans le tube non couvert d’huile indique une oxydation tandis qu’une coloration jaune dans les deux tubes indique une fermentation. d) Test d’hydrolyse de l’amidon (Lelliot et Stead, 1987). Une culture bactérienne de 24 heures est prélevée à l’aide d’une anse pour dessiner une figure serpentée sur le milieu d’amidon contenu dans une boîte de Pétri. Les boîtes de Pétri inoculées (une boîte par isolat) sont incubées pendant 3-4 jours à la température de 28-30°C. A la fin de la période d’incubation, les boîtes sont inondées avec l’iode de lugole. L’apparition d’une couleur jaunâtre autour ou sous la culture bactérienne indique une réaction positive. Par contre, si le milieu vire au bleu ou si on obtient une coloration rougeâtre, la réaction est négative, donc l’amidon n’est pas hydrolysé. e) Test d’hydrolyse de la gélatine (Fahy et Persley, 1983; Lelliot et Stead, 1987). Un tube à essai par isolat contenant le milieu gélatine est inoculé d’une culture bactérienne de 24 heures. Les tubes sont ensuite incubés à 28-30°C pendant 3-4 jours. Après incubation, les tubes sont placés à 6°C pendant 30 mn pour favoriser la gélification. Une réaction positive se traduit par une liquéfaction du milieu: on dit qu’il y a gélatinolyse. A l’issu des tests biochimiques, les souches dont les caractéristiques ont été conformes à celles de la référence de NCPPB 1150 sont utilisées pour les tests d’inoculation. f) Test d’hypersensibilité sur le tabac. Une suspension bactérienne de concentration 109 bactéries/ml est préparée à partir d’une culture de 24-48 heures. Cette suspension est ensuite injectée dans le mésophylle du limbe d’une feuille de tabac en introduisant l’aiguille d’une seringue hypodermique à 0,4 mm de diamètre environ dans la nervure de la feuille. La bactérie de référence et l’eau distillée stérile sont utilisées respectivement comme témoins positif et négatif. Les plants sont mis en incubation à la température de 28-30°C pendant 24, 48 et 72 heures. L’apparition d’un aspect vitreux suivi d’un brunissement des parties infiltrées indique une réaction positive. g) Test de pathogénie. La variété de riz TCS 10 sensible au flétrissement bactérien a été utilisée pour ce test. On prépare une suspension bactérienne de 109 bactéries/ml avec une culture de 48 heures. Des plantules âgées de 14 jours sont inoculées avec la suspension selon la méthode Kauffman et al. (1973). Deux semaines après inoculation, la bactérie est ré isolée des plantules flétries pour confirmer sa présence . La conservation des bactéries isolée se fait au congélateur sur le milieu de culture Yeast Glucose Chalk Agar medium (YGCA medium) dont la composition est la suivante : Yest extract (10 g) ; D-glucose (10g) ; CACO3 (10g) ; Agar (15g) et eau distillée (1000ml). INOCULATION DES PLANTULES DE RIZ Dispositif expérimental. Un dispositif Split plot à trois répétitions a été utilisé. Les deux souches de Xanthomonas oryzae pv. oryzae (XBFB et NCPPB1150) ont été affectées aux parcelles principales et les soixante neuf lignées aux parcelles secondaires. La souche NCPPB 1150 et l’eau distillée ont servi respectivement de témoins positif et négatif. Les variétés de riz TCS 10 sensible, et FKR 19 résistante ont servi de témoins. Culture du riz. Cinquante grains de chaque lignée préalablement désinfectés à l’eau chaude (53°C) pendant 30 mn ont été semés dans des pots (1,5 l) remplis de terreau stérilisé. Les pots sont placés sous serre à 30 °C avec une alternance de 12 heures de lumière et 12 heures d’obscurité. Inoculum et Inoculation. Des plantules âgées de 15 jours ont été inoculées suivant la méthode (Kauffman et al., 1973). Elle consiste d’abord à préparer une suspension bactérienne de 109 bactéries/ml. Les extrémités des feuilles sont sectionnées à l’aide d’une paire de ciseaux préalablement désinfectée à l’alcool et trempée dans la suspension bactérienne. Après inoculation, les plantules sont recouvertes d’un sachet plastique transparent et mises en incubation pendant 24 heures afin de favoriser l’infection. Evaluation des symptômes. Elle est réalisée sur la base de l’apparence générale des plantules d’une même répétition. L’échelle de notation utilisée est celle de Kauffman et al., (1973) (Tableau 3). Les observations ont été faites à 3, 7 et 14 jours après inoculation (JAI). Les moyennes des notes obtenues ont été regroupées dans des intervalles définis par l’échelle utilisée. Analyse des données. Le logiciel SAS a été utilisé pour l’analyse statistique des données. L’analyse de variance a porté sur la comparaison du pouvoir pathogène des souches de Xanthomonas oryzae pv. oryzae NCPBB 1150 et XBFB vis-à-vis des lignées. La comparaison des moyennes a été effectuée avec le test de ANOVA. Le logiciel Microsoft Excel 2000 a servi à la saisie des données. RESULTATS Les tests biochimiques et d’inoculation. Les souches isolées à Bagré ne sont pas fluorescentes mais elles présentent une coloration jaune caractéristique des xanthomonas. Elles présentent un test de GRAM (KOH) et d’ Oxydase négatifs. Elles sont toutes oxydatives et non fermentatives et ne réduisent pas non plus les nitrates. L’amidon et la gélatine ne sont pas hydrolysés par elles. Le test d’hypersensibilité sur le tabac est positif de même que celui de la pathogénie sur les plantules de riz. Elles ont les mêmes caractéristiques biochimiques que la souche de référence. Comportement des lignées vis-à-vis des souches bactériennes. L’analyse de variance (Tableau 4) indique qu’au seuil de 5%, les souches de Xanthomonas oryzae pv. oryzae diffèrent par leur pouvoir pathogène. Avec l’eau distillée stérile, aucune réaction de sensibilité n’a été observée. Les observations des plantules à 3, 7 et 14 jours après inoculation ont donné les résultats suivants (Tableau 4). A 3 JAI, quatre lignées à savoir WAS 122-IDSA-1-WAS-6-1 (V26), WAS 161-B-2-1 (V39), WAS 161-B-4-1 (V68) et le témoin sensible (TCS 10) ont présenté des symptômes. Les trois premières presentent ¼ d’infection foliaire à la souche XBFB et la TCS 10 à la souche de référene NCPPB 1150. Les autres lignées n’ont manifesté aucun symptôme aux deux souches bactériennes. A 7 JAI, trois groupes de lignées se sont illustrées:
Enfin, à 14 JAI, le comportement des lignées a été également evalué. Les moyennes obtenues et celles des 3 et 7 JAI sont présentées dans le Tableau 5. On note à partir des résultats de ce TABLEAU que la souche XBFB est plus agressive et plus virulente que la souche de référence NCPPB 1150. Sur 69 lignées testées, 16 lignées (02 intra, 14 interspécifiques) se sont révelées moyennement à hautement sensibles à la souche XBFB contre 08 lignées (01 intra-, 07 interspécifiques) à la souche NCPPB 1150. Par rapport à la résistance on observe que sur les 69 lignées testées 53 sont moyennement résistantes à résistantes à la souche XBFB contre 62 à la souche de référence. De plus, les lignéés V26, V41, V52, V61, V66 et V69 ont été sensibles à XBFB et résistantes à moyennement résistantes à NCPPB 1150. Seule la lignée V23 s’est révélée moyennement sensible à NCPPB 1150 et moyennement résistante à XBFB. DISCUSSIONS Analyse comparée du pouvoir pathogène des souches bactériennes. Le suivi de l’évolution de la maladie en fonction du temps montre que l’intensité des symptômes varie en fonction des lignées et des souches bactériennes de Xanthomonas oryzae pv. oryzae (Tableau 5). La souche isolée à Bagré (XBFB) s’est montrée plus agressive et plus virulente par rapport à la souche de référence (NCPPB 1150). La différence de comportement des deux souches testées vis-àvis des variétés serait liée à leur origine géographique. C’est du reste ce qu’affirment Horino et al. en 1981. Ces auteurs ont indiqué que les races de Xanthomonas oryzae pv. oryzae du Japon, région tempérée, diffèrent de celles des Philippines région tropicale humide. La souche de référence isolée au Bengladesh différerait par son agressivité et par sa virulence de la souche isolée au Burkina Faso (Bagré) à cause des conditions pédo-climatiques certes différentes. Analyse du niveau de résistance des lignées. Les lignées testées se sont comportées différemment par rapport aux deux souches (Tableau 5). Ces différences de comportement variétal vis - à – vis d’une souche donnée seraient liées aux caractéristiques intrinsèques à chaque variété (résistance ou sensibilité). Aldrick et al., (1973) ; Buddenhagen et Reddy, (1972) ; Ezuka et Horino (1974) cité par Mew et al., (1992) sont parvenus aux mêmes résultats en demontrant le phénomène d’interaction différentielle entre les variétés de riz et des souches bactériennes de Xanthomonas oryzae pv. oryzae. Kinoshita (1991) cité par Ahmed et al. (1997) signale que 18 gènes de résistance au flétrissement bactérien ont été recommandés. Plusieurs de ces gènes ont été incorporés dans des variétés de riz améliorées (Ogawa et al., 1991 cité par Mew et al. (1992) parmi lesquels, les gènes xa-5 et xa-7. Il est fort possible que certains de ces gènes soient incorporés dans les lignées résistantes rencontrées au cours de notre étude. L’un des multiples facteurs de la résistance provient de la progéniture. Mew et al. (1992) ont signalé que les cultivars IR utilisés aux Philippines sont modérement sensibles à une race de Xanthomonas oryzae pv. oryzae. Des résultats de leur étude, il ressort que la variété IR8 est sensible à 4 des 5 souches de Xanthomonas oryzae pv. oryzae utilisées alors que la DV85 possédant les gènes xa-1 et xa-5 a été résistante à toutes les 5 souches. Les 93,33% des lignées qui se sont revèlées sensibles dans notre étude ont un parent IR (TABLEAUX II et V), confirmant ainsi les résultats des études des auteurs cidessus mentionnés. Ainsi, si les cultivars IR sont moyennement sensibles à sensibles au flétrissement bactérien, la résistance des lignées testées serait liée en partie au cultivar TOG. La variété FKR 19, résistante aux souches XBFB et NCPPB 1150 peut être recommandée en riziculture de bas-fonds d’autant plus qu’elle est aussi résistante à la pyriculariose (Dembele, 2001). Mais d’après Ezuka et Sakaguchi (1978) cité par Mew et al. (1992), la variété Asakaze résistante au flétrissement bactérien a été sévèrement affectée par une souche virulente quelques années après son introduction aux champs. La variété FKR 19 pourrait également être sensible plus tard sur les sites de Bagré qui sont très infestés. En effet, l’apparition de nouvelles souches de la bactérie grâce au phénomène de mutation génétique peut rendre la FKR 19 vulnérable. Pour Reddy et Nayak (1984) cité par Ahmed et al. (1997), une réduction considérable de la sévérité du flétrissement bactérien est produite par le mélange de génotypes de riz résistants et sensibles. Le croisement entre la variété FKR 19, résistante et la TCS 10 sensible mais très productive pourrait être envisagé pour réduire la sévérité du flétrissement bactérien sur le périmètre de Bagré et améliorer la productivité du riz. CONCLUSION L’étude de la résistance variétale en conditions d’inoculation artificielles a permis d’identifier des lignées résistantes au flétrissement bactérien causé par Xanthomonas oryzae pv. oryzae. L’évaluation de ces lignées vis - à - vis de la bactériose au champ permettrait de proposer à la vulgarisation des lignées résistantes et plus promotteuses. Il serait intéressant d’ étudier la structure de la population du pathogène au Burkina Faso en utilisant les gènes Xa et xa disponibles dans les lignées isogéniques. De meme l’étude des gènes de résistance contenus dans les variétés évaluées ainsi que dans les variétés vulgarisées et en voie de vulgarisation au Burkina Faso, constitue une priorité de recherche sur le flétrissement bactérien. REMERCIEMENT Les auteurs remercient sincèrement le Centre du Riz pour l’Afrique (ADRAO), pour sa contribution financière et sans laquelle ces résultats ne s’auraient être atteints. Ces remerciements vont également au Prof. DAKUO Dona pour la lecture du manuscrit RÉFÉRENCES
©2007, African Crop Science Society The following images related to this document are available:Photo images[cs07020t2.jpg] [cs07020t5.jpg] [cs07020t4.jpg] [cs07020t1.jpg] [cs07020t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}