 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
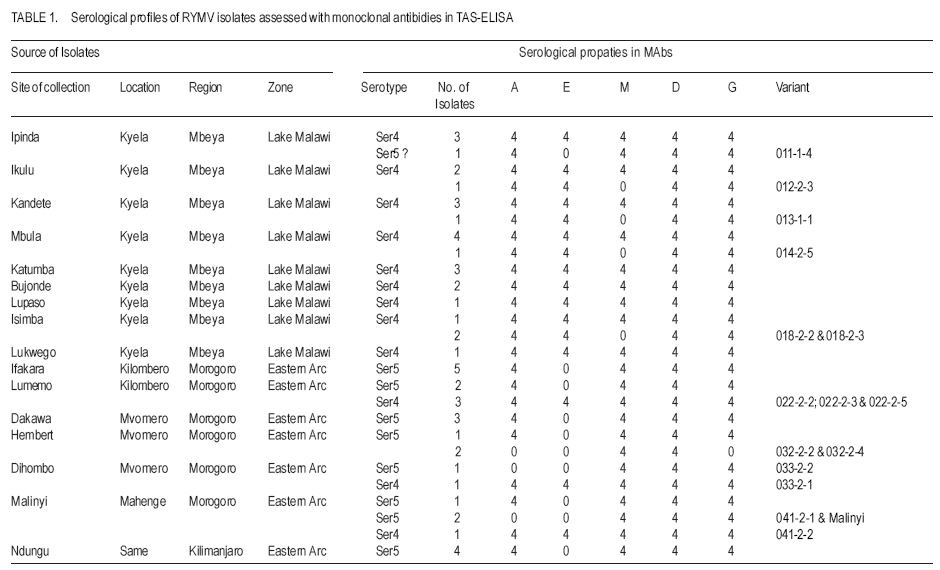
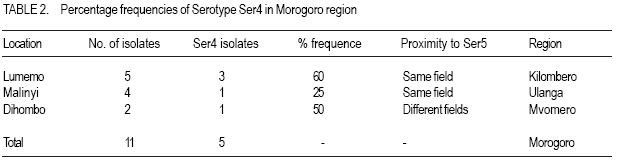
African Crop Science Journal, Vol. 15, No. 4, Dec, 2007, pp. 201-209 Distribution and Diversity of Local Strains of Rice Yellow Mottle Virus in Tanzania Z.L. Kanyeka, E. Sangu, D. Fargette1, A. Pinel-Galzi1 and E. Hérbrard1 Botany Department University of Dar es Salaam, Tanzania Code Number: cs07021 ABSTRACT Rice (Oryza sativa) is a key staple in Tanzania but its productivity is affected by disease infestation, primary among which is the rice yellow mottle virus (RYMV). Tanzania possesses high RYMV densities and several locations including the Eastern Arc Mountains provide suitable habitat fragmentation that facilitates the diversification process of the virus. However, the distribution pattern and diversity of the local strains of RYMV remain largely unknown and this hinders progress in designing interventions. Efforts were, therefore, made to study the pattern and distribution of RYMV. Collection of isolates was made in May; recovery of virus on a susceptible elite line viz. TXD 220-1-3-3-1 was in June-July and molecular analysis was done in September-December 2005. The sequences of the coat protein (CP) gene of 23 isolates, representative of the three strains of RYMV found in Tanzania, were selected from 52 isolates collected from three regions. The phylogenetic analyses of the CP sequences revealed high (14.8%) nucleotide divergence between strains. Strain S6 was the most divergent with an intra- strain nucleotide divergence of 8.8%; this was consistent with its wide distribution in the region and particularly the eastern part of the Tanzania. Isolates of strains S4 were recorded for the first time in the eastern Arc Mountain region. These formed a monophyletic group with the Lake Malawi strain S4 sub-lineage (percentage identity of 95.4 to 96.8%) and differed from the Lake Victoria strain S4 sub-lineage by 4.5%. Coexistence within the same field of S4, S5 and S6 isolates was observed in the eastern Arc Mountains region. Strong competition between strains of RYMV was suspected from the limited distribution and rarity of strain S5. This study confirm speculations that the Eastern Arc Mountain region is the centre of origin of RYMV. Thus, the high viral load in the Eastern Arc Mountain imply that it is a suitable location for strategic studies aimed at designing control interventions against RYMV. Key Words: Oryza sativa L., serological properties, strains, coat protein RÉSUMÉ Le riz (Oryza sativa) est un aliment de base en Tanzanie, mais sa productivité est significativement entamée par l’infestation des maladies, parmi laquelle est le virus jaune marbre de riz (RYMV). La Tanzanie possède les hautes densités de RYMV et plusieurs endroits y compris les Montagnes d’Arc d’Est fournissent de pochettes d’habitat qui facilite le processus de diversification du virus. Cependant, le modèle de distribution et la diversité des variétés locales de RYMV restent principalement inconnus et ceci freine le progrès dans la conception des interventions. Les efforts étaient, donc faits d’étudier la tendance et la distribution de RYMV. La collection des isolées a été faite au mois de mai ; i.e le rétablissement de virus sur une ligne d’élite susceptible. TXD 220-1-3-31 était fait au mois entre juin et juillet et l’analyse moléculaire a été faite entre septembre et décembre 2005. Les séquences de la protéine de manteau (PC) gênes de 23 isolées, été choisi de 52 isolées collectionnées dans trois régions. L’analyse phylogénétique des séquences de PC a révélé une grande (14.8%) divergence de nucléotides entre les variétés. S6 était la plus divergente avec une intra - divergence de nucléotide de 8,8% ; ceci était conforme à sa large distribution dans la région et notamment la partie Est de la Tanzanie. Les Isolées de S4 ont été identifiées pour la première fois dans la région de Montagne d’Arc d’Est. Cela a formé un groupe de monophylétique avec le sous lignage de S4 du lac Malawi (l’identité de pourcentage de 95,4 à 96,8%) et a différé du sous lignage de S4 du lac Victoria par 4,5%. La coexistence dans le même champ de S4, S5 et S6 a été observé dans la région de Montagnes d’Arc d’Est. La forte compétition entre les variantes de RYMV a été soupçonnée de par la distribution limitée de S5. Cette étude confirme des spéculations que la région de Montagne d’Arc d’est est le centre d’origine de RYMV. Ainsi, le haut chargement viral dans la Montagne d’Arc de l’Est implique que c’est un emplacement convenable pour les études stratégiques visées à concevoir d’interventions de contrôle contre RYMV. Mots Clés: Oryza sativa L., propriétés sérologiques, strains, protéine de manteau INTRODUCTION Rice yellow mottle virus (RYMV) of the genus Sobemovirus, is the most important plant virus of rice (Oryza sativa) in Africa. The virus is indigenous to the continent. Since its first observation on a local rice variety Sindano in a farmer’s field near Kisumu on the shores of Lake Victoria in Kenya in 1966 (Bakker, 1974), serious epidemics of RYMV have been reported in all rice ecological systems throughout Africa (Abo et al., 1998). Yield losses as high as 100% have often been reported, depending on the rice cultivars, time of infection, rice cropping systems and the isolates considered (Abo et al., 1998 ; Luzi-Kihupi et al., 2000). Rice yellow mottle virus is transmitted by beetles, artificial inoculation of healthy plants with virus infected sap, and through animals and human practices (Abo et al., 2000; Sarra and Peters 2003; Sarra et al., 2004), in addition to seed (Konaté et al., 2001). The virus has a narrow host range of plant species, restricted to cultivated (Oryza sativa and O.glaberrima) and wild (O. longistaminata and O.barthii) rice species, including a few wild grasses (Echinochloa colona and Panicum repens) (Bakker 1974; Konaté et al., 1997) Tanzania, one of the major producers and consumers of rice in sub-Saharan Africa, is severely affected by RYMV (Banwo et al., 2004). High virus incidence has been observed in nearly all rice producing areas, yet the major rice producing regions (Morogoro, Mbeya, Shinyanga and Mwanza) are the most seriously affected (Yamamoto et al., 1995). The virus poses a major threat to food security, in a country where about 60% of the population depend on rice as a staple food or as a source of income. This problem is complicated by the lack of effective control strategies that could help limit spread of virus. Tanzanian farmers are reluctant to adopt resistant rice cultivars, and to implement pest management strategies based on new farming practices which are partially effective to restrict virus spread. Seemingly, the high diversity of RYMV and the emergence of resistance - breaking RYMV isolates, further complicate the process of developing effective means of controlling the virus using resistant rice genotypes. Earlier studies have reported a high diversity of RYMV through serological and molecular typing. The diversity shows a pronounced and characteristic geographic structure with a strong relationship between the genetical and geographical distances (Fargette and Konaté, 2004; Traoré et al., 2005). Five serological profiles have been identified, 3 in West and Central Africa (Ser1, Ser2, Ser3) and the other 2 in East Africa (Ser4, Ser5). These serotypes were subdivided into 6 strains; 3 in West and Central Africa (S1, S2, and S3) and 3 in East Africa (S4, S5, S6). Serotype Ser5 was split into strains S5 and S6. Abubakar et al. (2003) revealed an exceptionally high diversity of RYMV in Tanzania. This was apparent through the number of strains, the level of divergence between the strains and the overall diversity per surface area (Abubakar et al., 2003). Seemingly, the Eastern Arc Mountain biodiversity hotspot harbours most of the strains found in East African, including the most variable strain S6 (Fargette and Konaté, 2004). Subsequently, it has been postulated that this area was the centre of origin of RYMV in Africa. From the Eastern Arc Mountains, the virus would disperse and differentiate gradually to Central and West Africa along an East to West transect (Traoré et al., 2005). According to this hypothesis of East to West dispersion across Africa, the absence of strain S4 in Eastern Arc Mountains was puzzling. Moreover, the narrow spatial localisation of most of Tanzanian strains and the high divergence between strains in Eastern Tanzania necessitated this study. Thus, the objectives of this study were to identify new strains of RYMV in Tanzania using molecular characteristics, and to assess their spatial distribution with emphasis on the processes of virus dispersion. MATERIALS AND METHODS Rice yellow mottle virus isolates were collected from 5 districts located in different regions. Kilimanjaro and Morogoro regions are traversed by the Eastern Arc Mountains, which run along the Tanzanian and Kenyan coasts from the border with Somalia in the north to Mozambique in the south. These chains of mountains (2000 m above sea level), together with the high altitude plateau (1200 m) separate the two regions from Mbeya in south-west Tanzania. Twenty five isolates were collected from 9 villages in Kyela district in the Mbeya region, near Lake Malawi (Lake Nyasa). Twenty-three isolates were collected from Kilombero valley at Ifakara and Lumemo (Kilombero district) and Malinyi (Ulanga district). Others were collected in Mvomero district at Dakawa, Hembert and Dihombo. The three districts are in Morogoro region. Kilombero valley is separated from Mvomero district by chains of mountains including the Udzungwa mountain. Other 4 isolates were collected from fields under rice irrigation schemes in Ndungu village, located at the foots of south Pare and Usambara mountains in Same district, Kilimanjaro region. The isolates were maintained on susceptible rice cultivars TXD 220 (Jaribu) at Botany Department, University of Dar es Salaam by artificial inoculation of 3 weeks old susceptible plants with inocula made by grinding infected rice leaves in PBST x 1 buffer pH 7.2. Symptomatic leaves were collected two weeks after inoculation and transported to Institut de Recherche pour le Development (IRD) France for serological and molecular analyses. Serological typing. Serological typing was performed using polyclonal and monoclonal antibodies. Virus was assessed by Direct double Antibody Sandwich Enzyme Linked Immunosorbent Assay (DAS-ELISA) method using a polyclonal antiserum raised against a Madagascar isolate (Ser4) according to Pinel et al. (2000) protocol. A sample of 0.2 g of leaf was ground in 2 ml PBST x 1 buffer pH 7.2 (1/10 w :v ratio) to obtain the leaf homogenate (antigen). Alongside 24 isolates representative of each site, an isolate from Madagascar and a healthy plant extract were tested, as positive and negative controls, respectively. Virus concentration in infected plant leaves was estimated with reference to a standard optical density reading of the positive control Madagascar isolate. The serological profile of the different isolates was determined using 5 monoclonal antibodies (MAbs A, D, E, G and M). Antibody M was nondiscriminative with the same affinity to each RYMV strain and was used as a reference to quantify the overall virus titer independently of the serological properties of the isolates. The MAbs were used in Triple-Antibody Sandwich-Enzyme Linked Immunosorbent Assay (TAS-ELISA) to test 52 isolates following the protocol of Pinel et al. (2000). Two polystyrene microplate wells were used to score the optical density values at 405 nm and the mean values were used for analysis. Molecular typing. Total RNA was extracted from frozen infected rice leaves using the Rneasy Plant Mini Kit (Qiagen), according to manufacturer’s instructions as it was done previously by Pinel et al. (2000). Slight modifications on the speed and time of centrifuging were made on the protocol to optimise the quality and quantity of the total RNA. A pair of primers, the sense primer 3’ M at 100 µM and antisence primer 5’ III at 100 µM were designed to transcribe and amplify the coat protein gene (Pinel et al., 2000). The sense primer 3’M was used in the Reverse Transcriptase Polymerase Chain Reaction (RT-PCR) to transcribe and amplify genome fragments containing the coat protein gene (nt 3447 to 4166) (Pinel et al., 2000). The PCR amplification of the RT-PCR products was done using the 2 primers (primer 3’ M 5’ III) under PCR conditions of denaturation at 94 °C for 5 minutes followed by 30 cycles of 3 minutes at 94 °C, 1 minute at 55 °C hybridisation, 1 minute at 72 °C elongation and final extension of 10 minutes at 72 °C. The mixture was stored at 4 °C. The PCR products were loaded and visualised by the electrophoresis of ethidium bromide stained 1% agarose gel in buffer TAE x 0.5 at 100V for 30 minutes. A nucleotide band of 720 bp was obtained. Coat protein gene sequencing. Sequencing of the coat protein gene was carried out with the Taq terminator sequencing Kit (Applied Biosystems) and analysed on an Applied Biosystem 373A sequencer (Pinel et al., 2000; Fargette et al., 2004). Two readings per base (3’to 5’ and 5’to 3’ directions) led to sequence accuracy of 99.9%. Sequences were assembled by Seqman (DNASTAR) and analysed using lasergene software by DNASTAR for Apple Macintosh computers (Madison). The coat protein gene sequences (720 to 723 bp) were found useful for assigning isolates to a particular strain, for estimating the inter and intra-strain diversity and for the assessment of the relationships between isolates. Phylogenetic analysis. The coat protein sequences of 16 isolates were aligned using Clustal V with default parameters. The alignments were checked by visual inspections and corrections made where necessary. The genetic distances between isolates were expressed in matrices of pairwise nucleotide divergence percentages (Nei and Kumar, 2000). Phylogenetic relationships between isolates were determined by the maximum parsimony method and a distance method. Nucleotide pairwise distances were corrected using Kimura’s two- parameter method and tree reconstructed by the neighbour-joining method, The phylogenetic analysis were performed using the Phylogenetic Analysis Using Parsimony (PAUP) computer programme version 4, (Swofford, 2000). Robustness of the nodes of the phylogenetic tree was assessed by 1000 bootstrap replicates of each data set. Isolates Tz2 and Tz3 were used as outgroups in rooting the phylogenetic tree. RESULTS Serological characteristics of the isolates. Two serological profiles were apparent using the discriminant monoclonal antibody panel. The first profile included all isolates collected from Kyela, Mbeya region (Lake Malawi) plus five other isolates from Morogoro region. They showed positive reaction with all the MAbs and were designated as serotype Ser4. The second serological profile included all isolates collected from Kilimanjaro region (Ndungu) and the remaining Morogoro isolates. These showed positive reaction with all MAbs except MAbE and were designated as serotype Ser5. Several variants of the two serological profiles (Ser4 & Ser5) were distinguished by their unusual reactions with different MAbs. Five isolates showed a negative reaction with MAb M, 5 isolates had a negative reaction with MAb A and 2 of the 5 isolates did not react with MAbs A and G, in addition to the discriminant negative reactions to MAb E. One isolate from Kyela also showed a negative reaction with Mab E. Table 1 shows response of 52 different isolates after tests with monoclonal antibodies in Triple Antibody Sandwich Enzyme-Linked immunosorbernt Assay (TAS ELISA). Positive reactions were scored from 1 to 4 according to intensity and negative reactions were scored 0. Table 2 gives the summary of the serological properties of the isolates and their respective sources of origin. The isolates belong to two serotypes Ser4 and Ser5. Ser4 includes all isolates from Kyela (Lake Malawi) and five isolates from Morogoro (Eastern Tanzania). Ser5 includes isolates from Kilimanjaro region and the remaining Morogoro isolates (Eastern Tanzania). All the five variants of Ser4 from Kyela had a negative reaction with MAb M. One variant showed a negative reaction with MAb E. Three variants of Ser5 from Malinyi (2) and Dihombo (1) in Morogoro region showed negative reaction with MAb A. Two variants of Ser5 from Hembert, Morogoro region, had a negative reaction with MAbs A and G. Malinyi has a large RYMV diversity with two different Serotypes (Ser4 & Ser5) and two variants of Ser5, followed by Lumemo and Dihombo, both with two different serotypes (Ser4 & Ser5). Ipinda, Ikuli, Kandete, Mbula and Isimba all share the same diversity with only one serotype and one type of variant.The isolates of a particular location generally share similar serological properties Ser4 isolates were found in Morogoro region for the first time, in three distantly located sites (Lumemo, Malinyi and Dihombo). The isolates were found within the same field or in different fields just a few meters apart but in the same location,with isolates of serotype Ser5. The frequencies of occurrence of serotype Ser4 isolates in the three sites are summarized in Table 2. However, strain S5 could not be distinguished from strain S6 with the serological typing. Molecular typing was necessary as it is more discriminative than serological typing. Identification of RYMV strains. The coat protein gene of the 16 isolates sequenced had the same length of 720 bp, except one isolate (22-2-4) which had a longer sequence of 723 bp. The longer sequence length was due to extra nucleotides (CGC) which codes for an extra amino acid arginine at position 60. This feature is typical of isolates of strain S5. The topology of the phylogenetic tree allowed the discrimination of three major strains with strong bootstrap support of bifurcating branches. The strains were exclusively found in East Africa region and were designated as S4, S5, and S6. No major variant was identified as there were no intermediates between the main clusters. Consistency between molecular typing and serological typing. There was a close relationship between serological and molecular typing although phylogenetic analyses were based on 720 characters (including 154 parsimony informative ones), whereas serological typing was based only on reactivity of four discriminating MAbs. Isolates of the same molecular strain had the same serological profile. This generalisation was not applicable to one isolate from Kyela, which showed no reaction with Mab E (a distinguishing reaction of Ser5 isolates), but was proved to belong to strain S4 with molecular typing. Serological typing sometimes gave ambiguous results and was less discriminating than molecular typing as it failed to distinguish strain S5 from strain S6. Distribution of RYMV strains. Assessment of the distribution of the 3 strains revealed that strain S4 occurs predominantly in Kyela district. It was also found in the 3 districts of Morogoro region (Mvomero, Kilombero and Ulanga). By contrast, strain S5 is restricted to a few sites only in Kilombero district. Strain S6 is widely spread in East Africa and occurs predominantly in all the 3 districts in Morogoro region and in Same district, Kilimanjaro region (Data not presented). The isolates of strain S4 in Morogoro were very closely related with percentage identity between 99.2 to 100. There was one exception of an isolate, which was more closely related to isolates from Kyela than to the rest of Morogoro. It differed from other Morogoro isolates by a percentage of 3.9 to 4.6%. S4 isolates in Morogoro were more closely related to the Lake Malawi strain with percentage identity between 95.4 to 96.8 than to the Lake Victoria strain (93.2 to 94.3%). Inter- and intra-strain molecular variability. The divergence among the 3 strains S4, S5 and S6 was high, with nucleotide sequence distance of the coat protein gene reaching up to 14.8% between the isolate from Lumemo (S4) and that from Ndungu (S6). The 2 variants of S4 in Lake Malawi strain and Lake Victoria differed between each other by nucleotide percentages ranging from 6.0 to 7.9. The Lake Malawi strain S4 was different from the Morogoro strain S5 by nucleotide percentage from 10.1 to 11.6%, and from strain S6 by 11.0 to 14.8. The Lake Victoria strain S4 differed from the Morogoro strain S5 by nucleotide percentage from 11.6 to 12.1, and from strain S6 by 10.3 to 12.5%. High nucleotide diversity was also observed between the Morogoro strain S5 and strain S6, both occurring primarily in the Eastern Arc Mountain, with divergence between 11.3 to 13.4%. The highest divergence (13.4%) recorded between these stains S5 and S6 was obtained with 2 isolates collected only 60 km apart. The intra-strain diversity was high among the isolates of strain S6 with a nucleotide divergence of 8.8%, as compared to 4.5% among isolates of Lake Malawi strain S4. Isolates of strain S5 and that of Lake Victoria strain S4 were relatively homogeneuous with nucleotide divergence < 2%. The high diversity among isolates of strain S6 was consistent with their wide range distribution within the surveyed area. Similarly, the expanded distribution of the Lake Malawi strain S4 has to some extent widened the divergence range among the isolates from 3.7 to 4.5%. Moreover, the narrow geographical distribution range of the strain S5 isolates was associated to a nucleotide divergence below 2%. Interestingly, in eastern Tanzania there was no relationship between geographic and genetic distances as highly divergent (10.1-11.6%) isolates were found to occur together within the same field. DISCUSSION New distributions of strain S4. Isolates of strain S4 have been reported to occur exclusively in East Africa region (Pinel et al., 2000). One variant includes all the Madagascar isolates and is distinguished from 2 other variants, the Lake Victoria S4 and the Lake Malawi S4 ones. Lake Victoria variant occurs predominantly in Mwanza, Shinyanga and Mara in the Tanzanian and Kenyan shores of Lake Victoria. Lake Malawi variant S4 was reported only in Mbeya region in the valleys opening to the Lake Malawi (Traoré et al., 2005). Our results show that S4 is also present in the 3 districts of Morogoro region, approximately 500 km from Mbeya region. The phylogenetic studies showed that these isolates are more closely related to isolates from Mbeya than to the Lake Victoria variant. These findings have never been reported before. Interestingly, similarities in gel diffusion tests between some isolates of the two regions have been reported. The distribution of S4 suggests that the RYMV crossed the extensive mountain barrier which separates Morogoro from Mbeya (Kipengere, Livingston plus the Eastern Arc Mountain Chain); and the mountain barrier that separates Kilombero district from Mvomero district (Udzungwa mountain) at least twice. No corridors can explain the mechanisms of the spread of the virus (Traore et al., 2005). Exchange of rice products for research and consumption between farmers in the two regions can not explain this, especially considering the fact that RYMV is not seed borne (Konaté et al., 2001). These results suggest that long distance spread (over several hundred kilometers) is possible, even when physical barriers are present. However, the pronunced geographical pattern of RYMV diversity indicates that such long distance spread must be rare events. More research work on RYMV ecology is necessary to understand the dispersion of this virus. The presence of Ser4 in the Eastern Arc Mountains reinforces the hypothesis that Eastern Arc mountain region is the centre of origin of RYMV. The dispersion processes from the centre of origin involved repeated movements of strain S4 to Lake Malawi and to Lake Victoria. Other movements may have reached Madagascar, and Central African countries. Analysis of isolates from Malawi and Mozambique would be useful to determine these routes of spread. Interestingly, the widespread strain S4 in Africa is not the the most common one in the centre of origin. Rare and highly localised strain S5. Despite extensive surveys conducted in the RYMV hotspot areas, the frequency of occurrence of strain S5 isolates have continuously been very low. The last collection of strain S5 isolates was made 5 years ago at Nazareti in the Kilombero valley. The current surveys were conducted in the same valley, at a nearby site, and collected only one isolate of strain S5 out of 23 isolates. This isolate was found together within the same field with several S4 and S6 isolates, the later in different fields a few kilometers away. Competition between strains have been demonstrated where two different strains coexist. Domination of strain S2 over S1 have been reported in West Africa, wherever S1/S2 coexist. Similar situation may have suppressed the occurrence and distribution of strain S5 and may explain the observed geographical pattern. Another limitation may come from the nonpersistent virus / vector relationship and massive inoculum extinction which would displace strain S5 isolates. However, isolates of this strain are infective both in the field and experimentally with moderate to severe symptom expression on susceptible host plants. Testing the fitness of strain S5 isolates may be worthy in elucidating the second postulation about massive extinction of inoculum. Our study has revealed the highest frequency of strain S6 in the Eastern Arc region, with 50% occurrence compared to strain S4 (41.67%) and strain S5 (8.33%). Similar results have also been reported by Banwo (2004) and Abubakar et al. (2003). Our results underline the high diversity of strain S6 with different variants. One branch groups together strain S6 isolates from Kilimanjaro and one Morogoro isolate (Tz 127). Another variant includes the remaining S6 isolates from Morogoro. This later variant is closely related to one of the Pemba isolates while the former relates to the other Pemba strain (S6). The two Pemba variants differed by 11.2% between each other (Traoré et al., 2005). Eastern Tanzania is known to be one of the global biodiversity hotspot zones. The high diversity of S6 isolates as well as the high genetic distances between S4, S5 and S6 strains support the hypothesis that primary diversification of RYMV took place there (Traoré et al., 2005). The presence of virus corridors may explain the successful spread of S6 isolates. Interestingly, despite its wide occurrence in the Eastern arc region, S6 apparently has not dispersed outside the Eastern Arc Mountains region. CONCLUSION Three RYMV strains (S4, S5, S6) occur in East Africa, particularly in Tanzania. Each strain has a specific and restricted geographical range. The new surveys gave valuable insights on their frequency and distribution. However, no new strains or major variants have been encountered. The findings suggest that the surveys conducted so far provided an exhaustive assessment of RYMV diversity. Then, analysis of the pattern of diversity should be conducted. In particular, coalescent analysis should reveal the major demographic events which shaped RYMV diversity. Accordingly, it would be interesting to test whether the lack of strain variants reflect massive extinction of the inoculum. Selection pressure may also counterselect intermediate strains. The extinction processess may be due to lack of infection of host, or lack of transmission by vector. The pronounced differences between strains may be due to independent colonisation events of different RYMV strains in different geographical areas. This idea was consistent with the pattern of node branching in the phylogenetic tree of all the RYMV strains in Africa both with the coat protein sequences and the sequences of the full genome. The study confirms the proposition that the Eastern Arc Mountain region in Tanzania is the centre of origin of the RYMV. Strain S4 is also found in the region thus reinforcing the proposition of the region as the center of origin of the virus. High genetic diversity of RYMV may be associated with emergence of resistance breaking isolates, which urgently calls for deliberate efforts to investigate this potential problem for future development of resistant cultivars to RYMV in the country. ACKNOWLEDGEMENT We are indebted to the to UDSM-Sida/SAREC Project of Enhancement of Molecular Biology and Biotechnology Research at the University of Dar es Salaam Project for financial support and to the Institut de Recherche pour le Development (IRD) for facilitating the molecular analyses of the isolates. REFERENCES
©2007, African Crop Science Society The following images related to this document are available:Photo images[cs07021t1.jpg] [cs07021t2.jpg] |
| |||||||||

{kind=link}
{kind=link}