 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
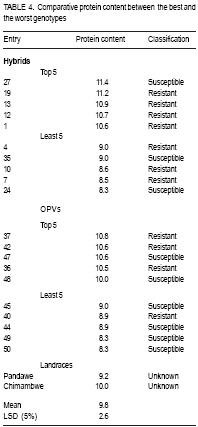
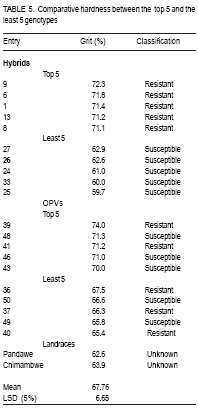
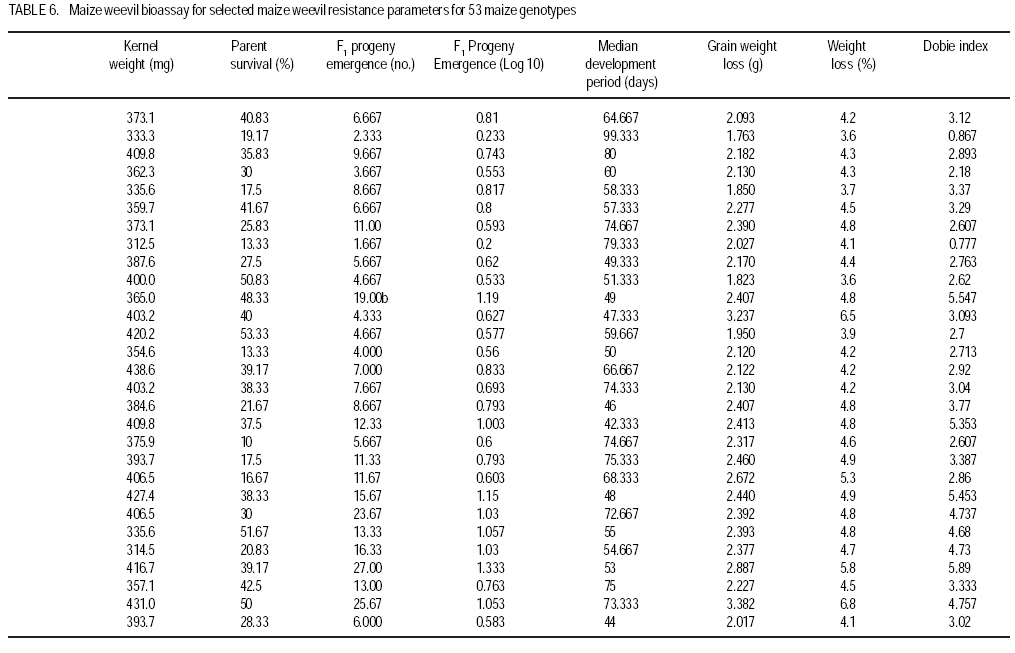
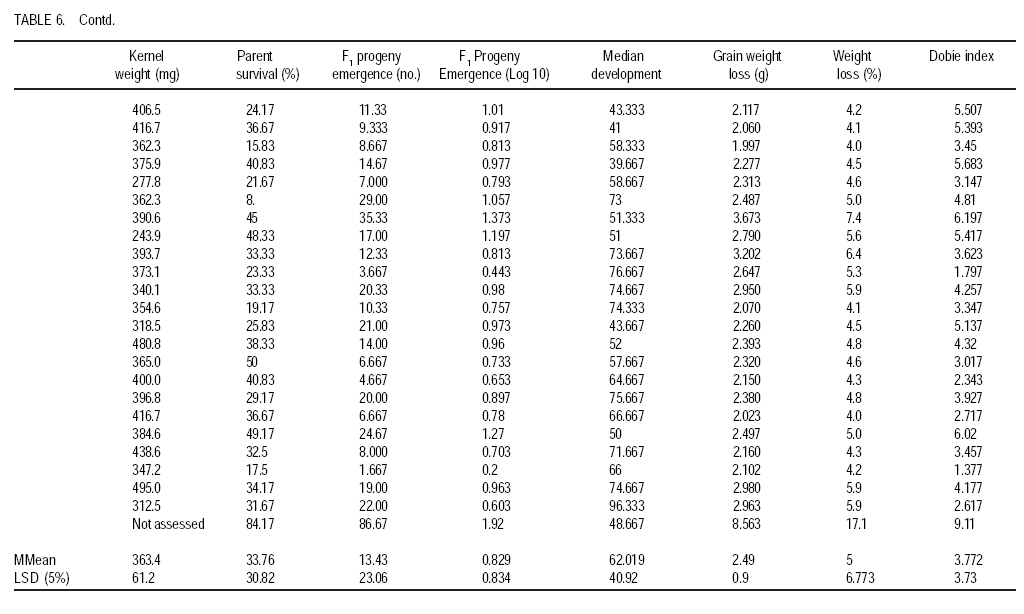
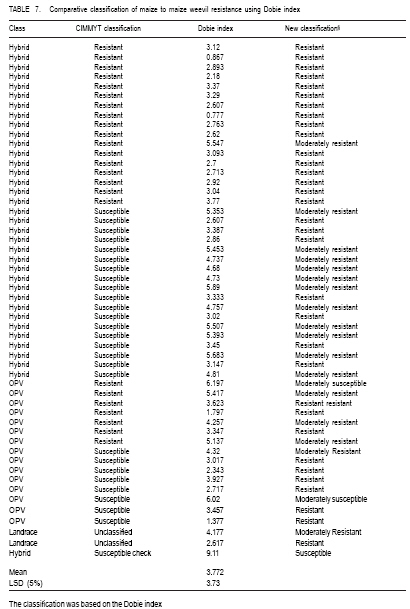
African Crop Science Journal, Vol. 17, No. 1, 2009, pp. 1-16 Comparative resistance of improved maize genotypes and landraces to maize weevil J. Siwale, K. Mbata1, J. Mcrobert2 and D. Lungu Department of Crop Science, School of Agricultural Sciences, University of Zambia, Lusaka, Zambia 1Department of Biological Sciences, School of Natural Sciences University of Zambia, Lusaka, Zambia 2International Centre for maize and wheat improvement (CIMMYT), Zimbabwe (Received 8 May, 2008; accepted 19 July, 2009) Code Number: cs09001 ABSTRACT A comparative study of maize (Zea mays L.) weevil (Sitophilus zeamais Motsch.) resistance in selected landraces and improved genotypes of maize, was conducted through a field experiment and laboratory assays. The maize grain characteristics that confer resistance to the maize weevil, Sitophilus zeamais Motschulsky, namely; grain hardness, weight and protein content, were compared in 50 improved maize genotypes developed by CIMMYT and two landraces (Cimambwe and Pandawe). The relative susceptibility of all maize experimental material to the weevil, were also compared using Dobie's susceptibility indices, grain weight loss and median development period. Genotypes were significantly different (p<0.05) for grain hardness but not for protein content. Genotypic differences in grain weight loss due to feeding by larval and adult S. zeamais were highly significant (P<0.001). The genotypes did not differ significantly (P>0.05) in the Dobie's indices of susceptibility but exhibited a wide index range (0.77 to 9.11). The landrace, Chimambwe, had the second highest Median Development Period, while Pandawe, had the heaviest kernels, though without noticeable advantage in resistance to the weevil. The Open Pollinated Varieties (OPVs) were not superior to hybrids according to the Dobie's index of susceptibility. The possibility of developing maize hybrids or OPVs that are as resistant to the maize weevil, S. zeamais, as or even better than landraces is discussed and recommendations made. Key Words: Open pollinated, Sitophilus zeamais, Zambia, Zea mays RÉSUMÉ Une étude comparative de résistance aux charançons de maïs (Zea mays L.) Curculionoidea (Sitophilus zeamais Motsch.) des cultivars traditionnels sélectionnés et les génotypes améliorés de maïs avait été réalisée à travers un champ expérimental ainsi que l'expérience et de laboratoire. Les caractéristiques de grains de maïs qui confèrent la résistance aux charançons de maïs à savoir, Sitophilus zeamais Motschulsky, dureté de grain; les poids et les contenu en protéines, avait été comparés pour 50 génotypes de maïs améliorés, développés par CIMMYT et deux cultivars traditionnels (Cimambwe et Pandawe). La relative sensibilité aux charançons de tout le matériel expérimental de maïs avaient également été comparé à l'aide de l'indices susceptibilité de Dobie, perte de poids de grain et la période médiane de développement. Les génotypes étaient significativement différents (p<0,05) pour la dureté de grains mais pas pour la teneur en protéine. Les différences génotypique en perte de poids grain en raison de la nutrition de larves et des adultes de S. zeamais. étaient hautement significative (P<0,001). Les génotypes n'ont pas différé significativement (P>0.05) dans les indices susceptibilité de Dobie de mais présentaient une gamme large d'index (0,77 à 9,11). Le cultivar traditionnel, Chimambwe, avait la deuxième plus longue période de développement médian, tandis que Pandawe avait les plus lourds amandes, pourtant sans avantage notable dans la résistance au charançon. Les variétés à pollinisation ouvert (OPVs) n'étaient pas supérieures aux hybrides conformément à l'index de susceptibilité de Dobie. La possibilité de développer des hybrides de maïs ou OPVs qui sont aussi résistants aux charançons de maïs, S. zeamais, comme ou même mieux que les cultivars traditionnels est discutée et les recommandations formulées. Mots Clés: Pollinisation ouverte, Sitophilus zeamais, Zambie, Zea mays INTRODUCTION Maize is the third most produced cereal after wheat and rice in the world. Its principle uses are human consumption and stock feed (Poehlman and Sleper, 1995). In Zambia, maize is the number one cereal in production followed by irrigated wheat (Zulu et al., 2000). The national yield of maize is low and is estimated at 1.5 tonnes per hectare (Pingali, 2001). Commercial yields of maize in the country are well below their potential which are estimated to be in the range of 8.5 t ha-1 for early maturing open pollinated varieties to 9.7 t ha-1 for intermediate to late maturing hybrids (Vivek et al., 2005). The low maize yields in Zambia and elsewhere in Africa are due to biotic and abiotic stresses on the crop. The International Maize and Wheat Improvement Centre (CIMMYT), in Harare, Zimbabwe, has classified maize production areas in Africa, into ecological zones and has documented the biotic and abiotic stresses that cause low maize yields in the zones. Zambia belongs to the mid-altitude, sub-tropical ecological zone within the sub-Saharan African region. Abiotic stresses depressing maize yield in this zone are low and declining soil fertility and drought. The biotic stresses include Gray Leaf Spot disease, streak virus disease, and damage by insect stem borers (Chilo and Sesamia spp.) and other insects including maize weevils. The latter infest maize while the crop is still in the field (Cardwell et al., 2000; Kim and Kossou, 2003; Pendleton et al., 2005; Asawalam and Hassanali, 2006) and destroy the crop during storage (Pingali, 2001). Despite the increased understanding of the inheritance of weevil resistance and of the resistance mechanisms in the maize grains, there has been very little application of this knowledge in maize breeding programmes (Dhliwayo and Pixley, 2002). Very little work has been done on breeding for maize weevil resistance in storage. In their study of maize weevil resistance involving two synthetic populations and four bi-parental populations, Dhliwayo and Pixley (2002) found that it was possible to improve weevil resistance in maize during storage using conventional breeding methods. The objectives of this study were to (i) compare the resistance of selected landrace and improved maize genotypes, to the maize weevil Sitophilus zeamais, and (ii) characterise grain traits that confer this resistance in the maize genotypes. MATERIAL AND METHODS Study area. A field experiment was set up at Golden Valley Agricultural Research Trust (GART) Station, 80 km north of Lusaka, Zambia, during the 2005 - 2006 growing seasons, to multiply the selected landrace and improved maize genotypes for laboratory weevil bioassays and biochemical tests. The experimental set-up was also to advance the F1 hybrids materials in the study to F2, the generation normally stored by farmers and which is what should therefore be resistant to the maize weevil damage (Munjoma, 2004). The Station is located at Latitude14o 40' South and Longitude 25o 01' East, and at 1140 m above sea level. The soil type is described as Makeni Series, which is a fine, mixed isohyperthemic ultic Paleustalf (Ti jmons, 1988). According to the World Reference Base (WRB), this soil is categorized as a Chromic luvisol (FAO, 1998). Field experiment. Thirty-five F1 hybrids and 15 Open Pollinated Varieties (OPVs) of maize obtained from CIMMYT and that had in previous studies been classified into various resistance group categories, were used in this study. Furthermore, two local varieties (landraces) from Mbala district of northern Zambia were also included in the experiment (Table 1a and Table 1b ). The two landraces are locally called Pandawe and Chimambwe. The description of these landraces is given in Table 2. A field experiment was laid out using a randomised complete block design with 3 replications. Each plot consisted of 4 rows measuring 5 m long. The inter- and intra-row spacing was 90 cm and 25 cm, respectively. Two maize seeds were planted per station. In addition to the standard crop management practices for maize, furadan was applied pre-plant in planting holes for the control of cutworms and stem borers; azodrin was applied against stalk borers, and confidor (imidacroprid) was applied at grain filling stage for the control of termites, which had already caused wide spread damage in the neighbouring maize trials. Methomyl, a carbamate, was applied against the armoured cricket Acanthoplus speiseri Brancsik, a pest of grain crops, common in the study area and elsewhere in central Zambia. Atrazine was applied for weed control and supplemented with hand weeding. For weevil evaluation, 12 plants were isolated from the two middle rows in each plot and upper ears covered with plastic bags to prevent pollination. Pollen was collected from the isolated plants, bulked and then used to pollinate the same plants (12 plants). Where plant emergence was poor, the number of isolated plants was less than 12. Laboratory assays. The maize weevils used in this study were collected from bags of weevil-infested maize, harvested from field trials at Mount Makulu Central Research Station, Chilanga, Zambia, during the previous growing season. The genotypes that were multiplied at GART were evaluated for maize weevil resistance by maize weevil bioassays using a modified Dobie's method (Dobie, 1977; Serratos et al., 1993). This was conducted at Mount Makulu Central Research Station. Three cobs of each genotype were hand-shelled and the grain packed into 5 x 8 polythene bags. The bags were closed with rubber bands and then stored in a deep freezer for one week to kill any previous infestation by insects, including adults, larvae or eggs (Kossou et al., 1993). The temperature in the freezer was minus -16 oC. Samples of 50 g of grains of each genotype were taken into new 350-ml plastic jars. These jars, purchased from Polymer Mouldings Limited in Lusaka, measured 11.7 cm in height and about 5.2 cm in diameter at the mouth. The tops of the lids of these jars were cut out, leaving only the screw-top rings. Forty unsexed weevils of mixed age, initially counted into vials with the help of pairs of tweezers and a Denominator Multiple-Tally tally counter (The Denominator Company, Inc. Woodbury, Connecticut, USA) were poured into each jar. To close the jars after the introduction of weevils, a piece of calico cotton cloth, 15 cm x 15 cm, was put on top of the jar and the ring of the lead screwed on to the jar over the cloth or the cloth was fastened to the jar with a rubber band. The cotton cloth was used to prevent the weevils from escaping and to provide ventilation. Grain of SC 513 maize variety from Seedco, an international seed company operating in Zambia, was included in the experiment as a susceptible variety check. This brought the total number of treatments to 53. Genotype entry numbers written on stickers were used to label the jars. The jars were placed in controlled temperature and relative humidity room and laid out in an RCBD with each shelf constituting a block. The temperature in the room was maintained at 29 ± 1 oC by a thermostat-controlled heater mounted on a wall. The relative humidity ranged from 43 to 60%; the humidity being provided by water placed in four troughs (Bekele and Hassanali, 2001). After an oviposition period of ten days, the adult (parents) maize weevils were removed from the samples by sieving with a Standard U.S.A. Testing Sieve set (VWR Scientific, West Chester, PA 19380,U.S.A.). The powder, where present, went through the No.8 (2.36 mm mesh opening) and the No. 18 (1.00 mm opening) and collected in the pan. Weevils went through the No 8 sieve and were collected on the No. 18 sieve; while the grain remained on the No. 8 sieve. Live and dead weevils were counted using tweezers and a tally counter. Tweezers were also used to probe immobile weevils to establish whether they were dead. Weevils, like some other beetles, tend to feign death when disturbed (Baker,2007). Sieving and checking for emergence of the F1 progeny started 3 weeks following the removal of the parents (Serratos et al., 1993). Sieving and counting the F1 progeny was done every 2 days (Derera et al., 2001) and the sieved insects were discarded. This interval of counts did not pose a risk of the F1 progeny laying eggs in the maize samples to produce the F2 generation, considering the fact that individuals of Sitophilus zeamais do not mate before they are three days old (Danho et al., 2002). Physical and biochemical parameters. Important maize kernel physical and biochemical parameters that have been reported to confer resistance to the maize weevil in the literature (Arnason et al., 1997) were analysed using appropriate methods. These parameters were grain hardness, protein content and kernel weight. Grain hardness. Grain hardness test was done by weighing a sample of 50 ± 0.1 g of maize kernels for each genotype. The sample was ground in a Retsh Laboratory Mill, Type ZM 1000 (GmbH & Co. KG 5657 HAAN 1, Germany). The grinding was done in two stages. During the first stage, the mill was set at 10,000 revolutions per minute (RPM) and 1 minute time setting for duration, with the sieve removed. This was done just to break the kernels into smaller fragments to make the next stage easier. The collected fragments were put back into the hopper and the number 11 sieve replaced. The speed and time setting was the same as above. The collected meal was put back in labelled plastic bags. The meal was then hand-sifted in a 0.5 mm DIN 4188 sieve (ANALYSENSIEB Retsck, W. Germany). The collected flour and retained grit were emptied in separate labelled 5 cm x 8 cm white plastic bags, and these were subsequently weighed and data recorded. The weight of the grit and flour were added together for each genotype to get the total weight, which was about the same as the original weight of the grain from where the flour and grit samples were derived. Grain hardness was expressed as percent grit of the total weight of the sample (grit plus flour after sieving a 50 ± 0.1 g ground maize sample). Thus, grit percentage was the proxy for grain hardness. Kernel weight. The number of kernels contained in a 50 ± 0.1 g grain sample of each genotype was determined and this number was divided into 50 g to obtain the weight per kernel. Protein content. Twenty-grammes samples of whole maize kernels were ground in a laboratory mill for each genotype. Protein content was determined using the Kjeldahl procedure. Dobie's Susceptibility Index. The Dobie index of susceptibility was used as the criterion to separate genotypes into different resistance groups (Dobie, 1977; Gudrups et al., 2001; Dhliwayo and Pixley, 2003). The index is given by the formula: I = 100 log e (no of adult weevil progeny emerged)/MDP. Where: I = Dobie's Susceptibility Index MDP = Median Development period, and this is the period (days) from the middle of the oviposition period to the middle of the emergence (i.e. 50 percent emergence) of the F1 progeny. Log e (sometimes written as log n) = the natural logarithm. The Dobie Index was then used to classify the genotypes into susceptibility groups following the scales used at CIMMYT in Zimbabwe (Pixley, 1997) which were as follows: Dobie index of < 4 was classified as resistant; Dobie index of 4.1 to 6.0 was moderately resistant; Dobie index of 6.1 to 8.0 was moderately susceptible; Dobie index of 8.1 to 10 was susceptible; and Dobie index of >10 was classified as highly susceptible. Statistical analyses. The Analysis of Variance (ANOVA) for all the measured parameters was done using the Mstat-C Programme (Freed et al., 1988). Total progeny emergence data were transformed to log base 10 before subjecting them to ANOVA (Dhliwayo and Pixley, 2003). Before conducting the log transformation, a value of 1 was added to all data points because of the presence of zeros in some data points in the data set. The mean separation, in cases where there were significant differences among treatments, was done using LSD (0.05) to facilitate the comparison of all pairs of treatment means (Montgomery, 2001). RESULTS Protein content. Genotypes were not significantly different (P>0.05) for protein content (Table 3). However, when the top 5 and least 5 genotypes in protein content in each group (hybrids and OPVs) were considered, it was found that resistant genotypes had a tendency of containing higher levels of protein than susceptible ones (Table 4). Grain hardness. Grain hardness showed discrimination among the 52 genotypes (P < 0.05). However, genotypes exhibited a higher or lower degree of hardness regardless of whether they were hybrids or OPVs. The two landraces were ranked low in hardness in relation with the top 5 hybrids and 5 OPVs. Separation of the genotypes into groups of the top 5 and the least 5 in grain hardness again showed that most of the harder genotypes were from the resistant class (Table 5). Parent survival. Table 6a and Table 6b presents the number of live weevils out of the 40 introduced in each sample. The difference is the number of weevils that were found dead in each incubation jar per genotype. The overall mean survival number for the parent weevils at the end of the oviposition period was 13.5, while the range was 4.0 to 33.7 weevils. Progeny emergence. Emergence of the F1 progeny was different among genotypes. Figure 1 shows an F1 adult progeny emerging from a kernel and an exit hole left by another F1 adult weevil. The total of all the F1 progeny adult weevils for each genotype is presented in Table 6a and Table 6b. The grand emergence mean was 13.43 and the range was 1.67 for entry number 8 and 50 to 86.67 weevils for the susceptible check. For the total weevil emergence data, the differences were highly significant (P<0.001) but for the transformed data, the differences were not significant (P>0.05). Grain weight loss. The greatest weight loss of 8.563 g occurred in entry 53, a susceptible check (Table 6a and Table 6b), while the lowest weight loss of 1.7 g was recorded in entry number 2. Dobie index of susceptibility. The Dobie Index of susceptibility ranged from 0.77 for entry numbers 8 and to 9.11 for the susceptible check (Table 6a and Table 6b). The trial mean was 3.77. However, the Dobie index re-classified the susceptible genotypes to be resistant or moderately resistant, except for the check which was susceptible (Table 7). DISCUSSION Protein content. Although genotypes were not statistically different for protein content (Table 1a and Table 1b), a closer look at the best 5 and worst 5 genotypes revealed a tendency for genotypes with higher protein content to be resistant (based on the classification of the genotypes done at CIMMYT, Zimbabwe). This is consistent with what other investigators have found (Derera et al., 2001; Dhliwayo and Pixley, 2003; Garcia-Lara et al., 2004). The fact that protein content did not have a definite relationship with physical resistance parameters in this study may indicate that there are other resistance factors in maize studied. Arnason et al. (1994; 1997) reported the presence of biochemical compounds, particularly ferulic acid in the kernels. Grain hardness. The differences among genotypes for grain hardness in this study were expected when there is a large number of genotypes in an experiment being evaluated. This is so because the genotypes had different grain textures. When grinding grain samples, genotypes with softer endosperms yielded more flour than those with harder endosperms. Grain hardness was closely related to maize weevil resistance (Table 5). These results are consisted with those of Leuschner et al (2000), who reported a distribution of larger numbers of Sitophilus oryzea progenies among genotypes of pearl millet (Pennisetum glaucum L) that had a higher proportion of soft endosperm. Parent survival. Parent weevil survival tended to be higher in susceptible than resistant genotypes. Thus, the susceptible check had the highest number, 33.7, of surviving parent weevils compared to the trial mean of 13.5. The larger number of parental survival generally leads to a larger number of eggs and ultimately the F1 progeny. The susceptible check yielded 86 F1 progeny compared to a grand mean of 13 and means of less than 1 in treatment 2 and 8 (Table 6a and Table 6b). Progeny emergence. Progeny emergence tended to be higher in susceptible genotypes than in resistant ones (Garcia-Lara et al., 2004). In this study, the susceptible check had the highest number of the total F1 emergence, numbering up to 87 weevils, against the experimental mean of 13 weevil emergencies. The total F1 progeny emergence may have been reduced in the whole experiment by mechanical disturbance of the samples through the action of sieving every 2 days (Ungunantwiwat and Mills, 1979). Median development period (MDP). The median development period in this study was very high, ranging from 39.667 in entry number 33, to 99.3 in entry number 2 (Table 6a and Table 6b). Other workers have reported lower ranges MDPs, for example, MDP ranges of about 4 to 40 days. The longer MDPs in this study may be attributed to the less than optimal relative humidity in the constant climate room. Grain weight loss. The highest loss again occurred in the susceptible check, in which 8.5 g were consumed against the experimental mean of 2.5 g. Grain weight loss values might have been higher than those obtained in this study if the weevils had been only young ones, 0 to 3 weeks. In his extensive experiments on the subject of maize weevil resistance, Dobie (1974; 1977) demonstrated that the fecundity and feeding of the maize weevils is highest when they are in the age range of 0 to 3 weeks after which there is a steady decline. Since the weevils that were used in this experiment were of unknown age, it is possible that some of them were older than the optimum age for feeding and reproduction. Dobie index of susceptibility. The range of values of indices obtained in this experiment ( 0.77 to 9.11) was lower than those obtained by other investigators. Arnason et al. (1994) obtained indices as high as 14 in susceptible varieties. One possible explanation is that the previous studies dealt with much more susceptible genotypes than in this experiment. Another cause could be the differences in moisture content. Most researchers infest their samples at about 14% moisture content. For instance, the maize samples that Arnason et al. (1994) used in Canada had moisture content ranging from 10.4 to 14.90%. The Dobie Index of Susceptibility from such maize samples then ranged from 0 for a resistant check to 15.2 for a susceptible check. The maize samples in the present study had moisture content of 10.5 to 12.5 percent. The Entomology Research Team at CIMMYT, Mexico, conducted a study to quantify the relationship between grain moisture content, kernel hardness, and resistance to S. zeamais and the larger grain borer Prostephanus trancatus (Coleoptera: Bostrichidae) (Bergvinson, 2001). They found that for grain moisture content below 12%, the resistant genotype, population 84, provided effective control for both insect species. However, once the moisture content reached 16%, the resistant (population 84) and susceptible (CML 244xCML349) entries showed similar damage levels. The age of weevils in the bioassay might also have contributed to the lower indices. Previous studies (Dobie, 1974) have shown that the fecundity of weevils is highest when they are 0 - 21 days old. The resistance/susceptibility of the genotypes in this study matched the classification of CIMMYT to a great extent in the case of hybrids, whereby 15 out of the 17 hybrids classified as resistant by CIMMYT were still found to be resistant However, there was no definite pattern for OPVS. This could be attributed to the variability in character of the OPVs. It was observed during the study that some OPVs had a mixture of normal white grain and some contained anthocyanin, and/or flint, as well as dent grain. The departure from the CIMMYT classification observed in some genotypes could be due to the effect of environment. Kim and Kossou (2003) studied the response and genetics of maize germplasm resistant to the maize weevil in Nigeria and found highly significant (P<0.01) data of crosses x location interactions for number of egg plugs, F1 weevils, damaged kernels and percent weevil survival. They concluded that the interactions indicated environmental effects on maize weevil resistance to weevils. Similarly, Duarte et al. (2005), in a study of nitrogen effects on grain quality of Brazilian maize genotypes, found that nitrogen application increased kernel hardness and decreased breakage susceptibility to a minor extent. However, according to these authors, genotype had a much larger influence on grain quality parameters than environment. ACKNOWLEDGEMENTS The authors thank Dr. J. MacRobert and Dr. C. Magorokosho (CIMMYT-Harare ) for supplying the seed for hybrid and open pollinated maize genotypes. Ms E. Namutowe and Ms. F.K Silutongwe, farmers of Mbala District in Zambia provided the seed for the land races. The Maize Research Team of the Zambia Agricultural Research Institute (ZARI) assisted in raising the maize materials for the study. Funding for this research was provided by the Rockefeller Foundation as a scholarship to the principal author to undertake a Masters degree course at the University of Zambia. REFERENCES
Copyright © 2009, African Crop Science Society The following images related to this document are available:Photo images[cs09001t3.jpg] [cs09001t5.jpg] [cs09001t1a.jpg] [cs09001f1.jpg] [cs09001t1b.jpg] [cs09001t4.jpg] [cs09001t6a.jpg] [cs09001t7.jpg] [cs09001t2.jpg] [cs09001t6b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}