 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
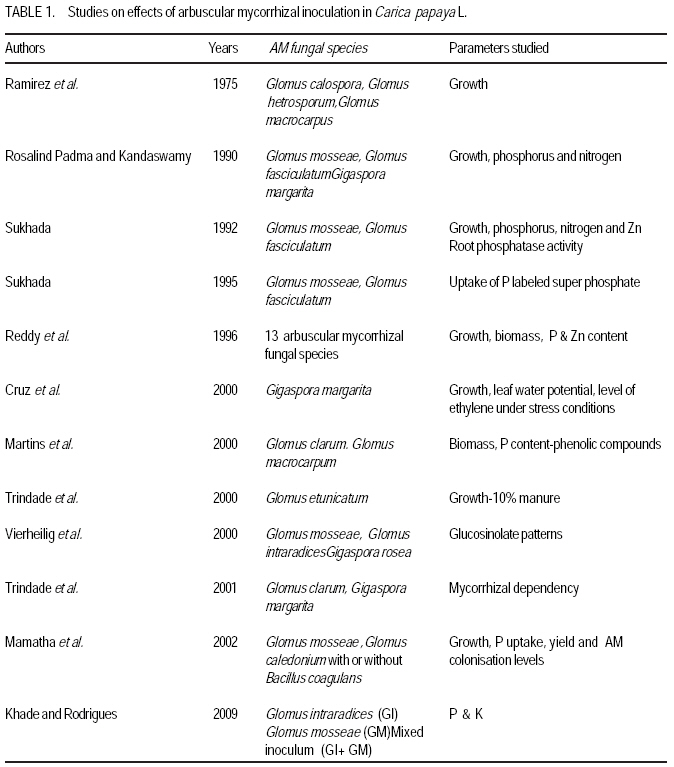
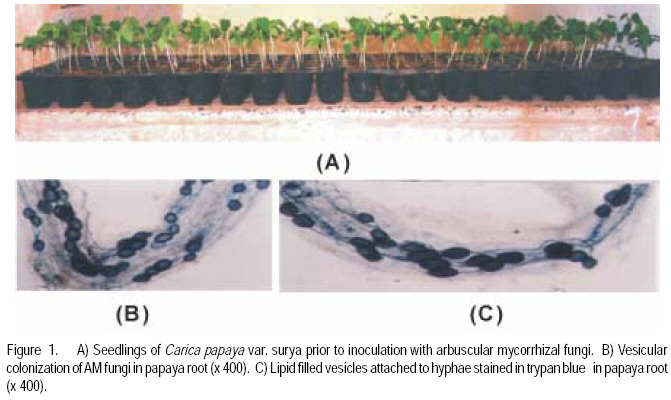
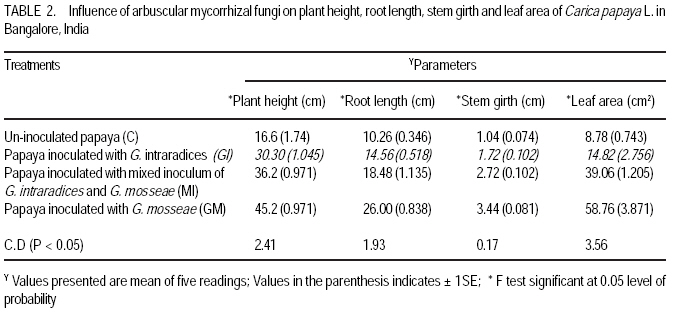
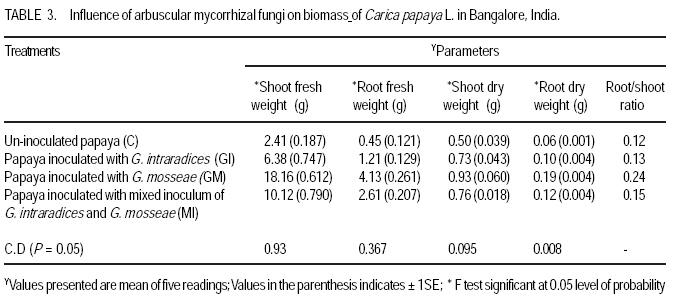
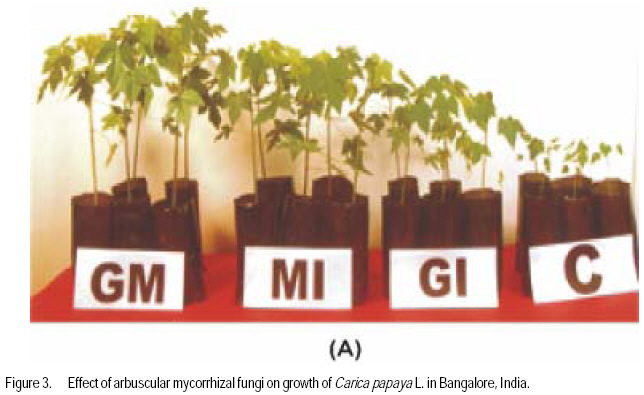
African Crop Science Journal, Vol. 17, No. 3, Sept. 2009, pp. 155-165 Studies on arbuscular mycorrhisation of papaya W. Sharda Khade and B. F. Rodrigues1 Vidhyanagar Colony, Carenzalem Post, Miramar, Panaji, Goa 403 002, India (Received 11 January, 2009; accepted 1 June, 2009) Code Number: cs09015 ABSTRACT Carica papaya L. is known to exhibit a strong growth response to colonisation by arbuscular mycorrhizal fungi; yet it is generally believed that mycorrhizal growth effects are primarily nutritionally mediated and are inversely related to improved soil fertility, especially available soil P, which affects the fungus symbiotic effectiveness. This study was, therefore, undertaken to evaluate the influence of selective arbuscular mycorrhizal fungi on vegetative growth of Carica papaya L. Studies were carried out in India on mycorrhization of Carica papaya var. Surya seedlings. The experiment comprised of un-inoculated seedlings, seedlings inoculated with Glomus intraradices, seedlings inoculated with Glomus mosseae and seedlings inoculated with mixed inoculum [Glomus intraradices + Glomus mosseae]. Inoculation with arbuscular mycorrhizal fungi significantly (P<0.05) increased all the growth parameters viz., plant height, stem girth, leaf area and root length. Mycorrhizal plants exhibited significant increase in fresh and dry weights of shoot and root systems. A higher root: shoot ratio was also observed in mycorrhizal plants than the control plants. Glomus mosseae was the most effective species of arbuscular mycorrhizal fungi, recording the highest root colonisation and exerting profound influence on growth of papaya. Key Words: Arbuscular mycorrhizal fungi, Glomus mosseae, Glomus intraradices RÉSUMÉ Carica papaya L.est caractérisé par une forte croissance en réponse à la colonisation des champignons mycorrhiziens arbusculaires ; de même il est généralement admis que les effets de la croissance des champignons mycorhiziens jouent un rôle principalement nutritionnel et sont inversement liés à l'amélioration de la fertilité du sol, spécialement à la disponibilité en P dans le sol, ce qui affecte l'efficacité symbiotique du champignon. Cette étude a été entreprise pour évaluer l'influence de la sélectivité des champignons mycorhiziens à arbuscules sur la croissance végétative de Carica papaya L. Des études ont été faites en Inde sur la mycorhization des plantes de Carica papaya var.Syria. L'expérience était composée de plantes non inoculés, de plantes inoculés avec Glomus intraradices et des plantes avec l'inoculum mixte (Glomus intraradices + Glomus mosseae). L'inoculation par des champignons mycorhiziens à arbuscules a montré une augmentation significative (P<0.05) de tous les paramètres de croissance à savoir la hauteur de plantes, le diamètre de la tige, la surface foliaire et la longueur des racines. Les plantes avec mycorhizes ont montré une augmentation significative de la surface foliaire et de la longueur des racines. Egalement, le rapport pousse-racines a été plus élevé chez les plantes avec mycorhizes que celles sans mycorhizes. Glomus mosseae a été l'espèce la plus efficace de mycorhizes à arbuscules, ayant enregistré un grand nombre de racines colonisées et une grande influence sur la croissance du papaya. Mots Clés: Champignons à mycorhizes arbusculaires, Glomus mosseae, Glomus intraradices INTRODUCTION Carica papaya L. is known to exhibit a strong growth response to colonisation by arbuscular mycorrhizal fungi (Ramirez, 1975; Sukhada, 1989). It is well documented that mycorrhizal growth effects are primarily, nutritionally mediated and are inversely related to improved soil fertility, especially available soil P, which affects the fungus symbiotic effectiveness (Rosalind Padma and Kandaswamy, 1990; Sukhada, 1992). Recent studies have confirmed these findings and have demonstrated the importance of arbuscular mycorrhizal symbiosis to papaya in low fertility soils (Martins, 2000). Also, they have elucidated their mycorrhizal status with respect to glucosinolate patterns (Vierheilig et al., 2000) and investigated the effect of arbuscular mycorrhizal fungi on leaf water potential and ethylene levels under stressed conditions (Cruz, 2000). Mamatha et al. (2002) have observed that papaya trees respond to inoculation with efficient arbuscular mycorrhizal fungi reducing the need for P fertilisation at least until the start of production. Kuwada et al. (2006) also reported that root colonisation of papaya was markedly stimulated by the application of red and green algal extracts. On the other hand, Khade and Rodrigues (2009) reported increased in elemental P and K in papaya plants inoculated with arbuscular mycorrhizal fungi. Similar studies carried by the earlier workers on papaya are summarised in Table 1. The widespread occurrence of arbuscular mycorrhizal fungi in soils throughout tropical and temperate regions has sometimes led to the notion that inoculation of soils with arbuscular mycorrhizal fungi is not essential. However, inoculation is necessary where the fungi have been eliminated or their populations are reduced by pesticide application, fumigation, erosion, or other forms of soil disturbance. In some instances, indigenous arbuscular mycorrhizal fungi may either express their symbiotic effectiveness after a prolonged lag phase, or their inherent effectiveness may be too low, and thus this needs to be preempted by more aggressive, highly effective arbuscular mycorrhizal inocula (Habte and Fox 1989). Arbuscular mycorrhizal fungi play crucial roles in agricultural situations, including: (i) agricultural systems in which the high P-fixing capacities of soils and the unavailability or high cost of P fertiliser limits crop production, (ii) situations in which it is essential to reduce soil fertiliser application rates significantly because of environmental concerns, and (iii) situations in which phosphate rock is readily available and used instead of super phosphate (Habte, 2000). In the view of tremendous potential for arbuscular mycorrhizal fungi in maintaining sustainability of agro-based ecosystem, studies on the effect of arbuscular mycorrhizal fungi on growth of papaya would provide information essential to attempt to use these fungi in environmental conservation (Allen, 1991), biotechnology (Mulongoy et al., 1992) or in sustainable agriculture (Bethlenfalvavy and Linderman, 1992). This study was, therefore, undertaken to evaluate the influence of selective arbuscular mycorrhizal fungi on vegetative growth of Carica papaya L. MATERIALS AND METHODSPlant material. Seeds of Carica papaya var. Surya were procured from Indian Institute of Horticultural Research (IIHR), Bangalore. Surya is gynodioecious, high yielding (50-65 kg plant-1) variety and is a progeny from the cross between Sunrise Solo and Pink Flesh Sweet released by IIHR, Bangalore. To date no literature exits on its mycorrhization. Fungus material. Sand based pure cultures of Glomus mosseae (Nicol. & Gerd.) Gerd. & Trappe and G. intraradices Schenck & Smith containing extramatrical chlamydospores, colonised root segments containing mycelium and vesicles were obtained from The Energy Research Institute (TERI), New Delhi. The inoculum of each culture consisted of 30 spores 2 g-1 moist soil. Growing conditions. The experiment was conducted for four months (May 2002- August 2002) in a poly-house in agriculture farm located at Mapusa, North Goa. The relative humidity during the study period ranged from 84 to 93%. Maximum and minimum temperatures ranged from 31.6 to 29.3 0C and 25.4 to 24.10C, respectively. Throughout the experimental period, papaya plants were watered thrice a week and fertilised with Hoagland's nutrient solution without phosphorus (Hoagland and Arnon, 1935), at an interval of fifteen days. Raising of seedlings. Seeds of papaya were sown in plastic trays with sterilised nursery soil (sand soil mixture, 1:1). Garden soil was sterilised for two hours daily at 15lb pressure for three consecutive days to eliminate naturally occurring endophytes and other contaminants. The soil used for the experiment was low in available phosphorus (6 kg ha-1) (Brays method, 1945) at pH 6.1 and electrical conductivity 0.06 m mhos cm-1 (conductivity meter, CM-180, Elico). Total nitrogen (Jackson, 1971) and exchangeable potassium (Hanway and Heidel, 1952) was 0.4% and 80 kg ha-1, respectively. Organic carbon content (Walkley and Black method, 1934) was also low (0.42%). Available Zn2+, Cu2+, Fe2+ and Mn2+ (Lindsay and Norvell, 1978) concentrations were 2.67, 3.84, 2.63, and 19.5 ppm, respectively. Seedlings were maintained in plastic trays for one month. Inoculation with arbuscular mycorrhizal fungi. Seedlings of uniform length (10 cm) were selected for inoculation with arbuscular mycorrhizal fungi at the end of one month (Fig. 1A). The experiment comprised of four treatments and each treatment had fifteen replicates. The treatments were: Treatment 1 (C) Un-inoculated Carica papaya var. Surya seedlings; Treatment 2 (GI) - Carica papaya var. Surya seedlings inoculated with G. intraradices; Treatment 3 (GM)- Carica papaya var. Surya seedlings; inoculated with G. mosseae; and Treatment 4 (GI + GM) - Carica papaya var. Surya seedlings inoculated with mixed inoculum G. intraradices + G. mosseae. Nursery polythene bags of 0.5 kg capacity were filled with sterilised soil up to 75% of their volume. A small pit was made into soil and 5 g of inoculum was placed at a depth of 5 cm. Papaya seedling was then placed in this pit and layered with sterilised soil. One seedling was planted per bag. In the case of GI + GM inoculation, equal quantity (75 g each) of inoculum of G. intraradices Schenck & Smith and G. mosseae were mixed thoroughly and then 5 g of inoculum added to each nursery bag. Control plants were inoculated with autoclaved inoculum. The plants were maintained for one and half months (45 days) in sterilised soil. Transplantation of seedlings in unsterilised soil. After 2.5 months of growth (76 days), papaya seedlings were transferred to nursery polythene bags of 3 kg capacity, containing unsterilised nursery soil with low nutrient status as mentioned earlier. The plants were further maintained for a period of 45 days. The experiment was terminated at end of 123 days and papaya plants were subjected to analysis for various parameters as follows. Estimation of root colonisation of arbuscular mycorrhizal fungi. Three plants per treatment were randomly selected for detecting the presence of arbuscular mycorrhizal colonisation. Papaya roots were thoroughly washed with tap-water, cleared with 10% KOH, acidified with 1N HCl and then stained with 0.05% trypan blue (Phillips and Hayman, 1970). The degree of root colonisation was estimated using the slide method (Giovannetti and Mosse, 1980) and the values were expressed as percentages. Plant growth measurements. Parameters viz., plant height, leaf area, stem girth, root length, fresh weights of root and shoot system, and dry weights of root and shoot systems were evaluated. There were five replicates for each treatments. Plant height was measured from the surface of soil to the highest point of the plant. Also, leaf lamina area was measured using a non-portable leaf area meter (Systronic 21). Stem girth was measured 2 cm above the surface of soil. For fresh weights, plants were carefully removed from the polyethylene bags and the root and shoot portions were separated. Roots were thoroughly washed with tap-water to remove soil debris and then dried using blotting paper. Root lengths were recorded. The root and shoot systems were weighed separately. Further more, root and shoot potions were oven-dried separately at 70 0C for 72 hr and dry weights recorded. RESULTS Microscopic observations revealed that in all the treatments, roots of papaya exhibited colonisation by arbuscular mycorrhizal fungi. Extensive mycorrhizal colonisation was observed in inoculated plants compared to the control. However, colonisation was confined only on first and second order roots; the first order being absorbing rootlets, according to the morphometric classification of roots (Fitter, 1982). In the present study, it was observed that the visible colonisation by arbuscular mycorrhizal fungi was confined to vesicles with dense lipid droplets and the adjacent hyphae which were, in most instances, weakly stained (Fig. 1 B and C). At the harvest, maximum mean total root colonisation was recorded in plants inoculated with G. mosseae (84%), followed by mixed inoculum (GI + GM) (72%) and G. intraradices (58%) with values given in parenthesis. The unsterilised nursery soil contained enough propagules of mycorrhizal fungi, which developed a natural colonisation level of 22% in un-inoculated control after transplanting. Data on the growth parameters of papaya plants are presented in Tables 2 and 3. Inoculation with arbuscular mycorrhizal significantly (P<0.05) increased plant height, stem girth, and leaf area (Fig. 2A), and root length (Fig. 2B) over the to control. Mycorrhizal plants exhibited significant (P<0.05) in fresh weight increases of shoot and root systems (Table 3). Also, significant (P<0.05) increases in dry weights of shoot and root system of mycorrhizal plants were recorded over the control plants. Additionally, comparatively higher root: shoot ratio was observed in mycorrhizal plants than control plants (Table 3). In general, inoculated plants exhibited increased growth over the control plants. However, there was wide variation in the growth promoting efficiency of arbuscular mycorrhizal fungi (either singly or in combination). Glomus mosseae was the most effective species of arbuscular mycorrhizal fungi, exerting profound influence growth of Carica papaya var., Surya ; followed by mixed inoculum (GI +GM) and G. intraradices (Fig. 3). DISCUSSION The study clearly demonstrates the benefits of inoculation with arbuscular mycorrhizal fungi during early developmental stages of papaya grown in unsterelised soil under low levels of phosphorus. This potential of arbuscular mycorrhizal fungi to increase plant growth under low soil P has been well documented by several workers (Covey et al., 1981; Gnekow and Marschner, 1989; Graham et al., 1996). Generally, inoculation studies with soil having indigenous mycorrhizal population are problematic since the colonisation caused by the introduced fungi cannot be conclusively attributed to the inoculants. This is especially so if there is doubt that the inoculum dose and infectivity have been significantly high (Niemi and Vestberg, 1992). In the present study, however, each inoculum dose per plant consisted of 75 spores, a substantial amount of colonised root fragments plus an unknown amount of external hyphae. Thus, root colonisation was not limited by lack of infective propagules. The fact that inoculation with arbuscular mycorrhizal fungi increased the overall plant growth more than un-inoculated control, indicated that the inoculation of papaya was successful under the given experimental conditions. Cruz et al. (2000) also employed a similar number of spores of arbuscular mycorrhizal fungi for successful mycorrhization of papaya (var. Solo) plants in a controlled experiment. The results obtained from the present study revealed differences among arbuscular mycorrhizal fungi in their ability to colonise the roots and influence growth in papaya plants. These inter-specific variation in the efficiency or growth promoting abilities of arbuscular mycorrhizal fungi could be attributed to the mechanism of mycorrhizal colonisation development (Sanders et al., 1977) or interaction between mycosymbionts and soil environment (Mosse, 1973). Of the three mycorrhizal treatments tested for their efficiency, inoculation with G. mosseae significantly (P<0.05) increased all the study parameters ,followed by mixed inoculum, and in turn inoculation with G. intraradices. Thus, these results substantiate the hypothesis that not all combinations of host and endophyte have similar growth stimulating effects (Hayman, 1980), and this may be interpreted as a kind of functional host specificity or compatibility. Sukhada (1989) also reported G. mosseae to be more beneficial compared to G. fasiculatum in a glass house experiment where papaya (cv. Coorg Honey Dew) seedlings were inoculated in sterilised acidic soil and then transplanted to unsterile soil with varying levels of phosphorus. Reddy et al. (1996) screened the efficiency of 13 arbuscular mycorrhizal fungi in papaya plants grown in acidic unsterile soil amended with two percent organic matter, and reported that G. mosseae (ICRSAT) was the most efficient fungus for improving growth and nutrition of papaya. Although it has been shown repeatedly that the amount of colonisation by arbuscular mycorrhizal fungi is not always correlated with efficiency (Yosef et al., 1984; Declerck et al., 1995), the present study appears to demonstrate a relationship between extent of colonisation and mycorrhizal efficiency. Plants inoculated with G. mosseae recorded the highest total root colonisation and maximum plant growth, followed by mixed inoculum (GI + GM) and G. intraradices. These results are in agreement with earlier findings of Sanders et al. (1977), who reported that arbuscular mycorrhizal fungi which are more effective in increasing growth, colonise plants more rapidly and extensively. Similarly, Miller et al. (1989) revealed that apple plants inoculated with G. mosseae and G. mosseae + G. macrocarpum recorded higher root colonisation and growth rates than plants inoculated with G. macrocarpum only. Another factor undoubtedly important in explaining higher symbiotic efficiency of G. mosseae, but not estimated in this study, could be the amount or intensity of hyphal distribution in the soil. If the extramatrical hyphae are correlated with intramatrical hyphae (Tisdall and Oades, 1979), then the differences in the fungal species may be due to differences in the volume of soil explored (Miller et al., 1985). Control plants of papaya were colonised by indigenous mycorrhizal and recorded lower root colonisation than the plants inoculated with selected mycorrhizal fungi; this is in agreement with the findings Vestberg (1992) and de Silva et al. (1996) in strawberry. Low levels of root colonisation in control plants indicated that the nursery soil had few indigenous fungi at low inoculum density (Ragupathy and Mahadevan, 1995) and that these indigenous fungi may have influenced the growth of control plants (de Silva et al., 1996) after transplantation in unsterile soil. However, inoculated plants had an added advantage over the indigenously colonised plants since they were colonised before transplanting (de Silva et al., 1996). In this study, greater shoot length of inoculated plants was accompanied by larger leaf area, increased stem girth and root length compared to control plants in unsterile acidic soil, with low phosphorus. This is in agreement with the work of Plenchette et al. (1981) and Reddy et al. (1996) who reported similar growth response in apple and papaya inoculated with arbuscular mycorrhizal fungi grown in unsterile acidic soil with similarly low levels of phosphorus. The study also supports the findings of Onkarraya and Mohandas (1993) that presented improved height and root length of mycorrhizal Citrus under similar set of experimental conditions. Ramirez et al. (1975) reported increased shoot length of papaya plants inoculated with Scutellospora calospora and G. macrocarpum, and the plants were taller than plants inoculated with S. heterogama, and the uninoculated controls after 40 days of sowing. Sukhada (1989; 1992) also reported increased in plant height of Carica papaya cv. Honey Dew inoculated with G. mosseae and G. fasiculatum under varying levels of phosphorus, after 5 months of growth in a glass house. Rosalind Padma and Kandaswamy (1990) recorded 25.47% increase in plant height of papaya after 90 days of growth by application of 75% of recommended dose of phosphorus along with mixed inoculum (G. mosseae + G. fasciculatum + Gigaspora margarita). Thus, the increased heights in mycorrhizal plants could be due to the fact that mycorrhiza can cause earlier resumption of shoot apical growth (Azcoñ- Aguilar and Barea, 1997). Arbuscular mycorrhizal fungi are known to induce morphological modifications in the host plant root system and a more branched root system has been observed in mycorrhizal plants of different herbaceous and woody species (Berta et al., 1995). In the present study, mycorrhizal inoculation of papaya increased the length of the taproot and the lateral roots of first and second order. This is in agreement with Bhattacharaya et al. (2002) who studied the changes in root development patterns of sweet orange inoculated with G. mosseae, G. fasciculatum and Gigaspora margarita under acidic soil with low levels of phosphorus. Papaya (var. Surya ) plants inoculated with mycorrhizal fungi also recorded increased biomass compared to uninoculated controls. The most significant response was exhibited in plants inoculated with G. mosseae. These observations support the earlier findings of Reddy et al. (1996) who reported that G. mosseae (ICRISAT) was the most efficient fungus for improving plant biomass of papaya plants. Jaizme-Vega and Azcoñ (1995) reported G. fasiculatum to be the most efficient fungus in improving growth of papaya, pineapple and banana under green house and field conditions. Rosalind Padma and Kandaswamy (1990) reported increase in plant biomass of papaya after 90 days of growth by application of 75% of recommended dose of phosphorus along with mixed inoculum (G. mosseae + G. fasciculatum + Gigaspora margarita) than control plants. In the present study, the dry weights of inoculated plants were significantly greater than control plants, in soil of low P levels with acidic pH. In a glass house experiment with soil of alkaline pH and varying P levels, Menge et al. (1978) reported significantly greater dry weights in mycorrhizal citrange and sour orange seedlings. In their study, maximum increase in dry weights were recorded at low levels of P viz., 0 and 6 ppm for mycorrhizal citrange (746 and 300%) and 947 and 1089% for sour orange. Similarly, Sukhada (1997) reported increased accumulation of dry matter in mycorrhizal papaya (cv. Honey Dew) in a glass house experiment, where seeds were directly sown in soil treated with arbuscular mycorrhizal fungi. She reported that G. mosseae was more efficient than G.fasiculatum in improving plant dry matter. In an earlier experiment comparing the efficacy of arbuscular mycorrhizal fungi under sterile and unsterile soil, Sukhada et al. (1995) reported that papaya plants inoculated with G. mosseae exhibited an increased dry matter (26.6%) than G. fasiculatum (4.3%) under sterile conditions. However, in unsterile soil, G. fasciculatum recorded higher accumulation of dry matter (29.5%) compared to G. mosseae (26.3%). de Silva et al. (1996) also recorded 70% more dry matter in mycorrhizal strawberry relative to control plants. Usually, mycorrhizal plants are known to exhibit lower root/shoot ratio than control plants in response to greater increment of shoot mass relative to root mass (Onkarayya and Mohandas, 1993). Estrada Luna et al. (2000) reported comparable values between the root/shoot ratios of mycorrhizal versus non-mycorrhizal micro-propagated guava plantlets after 18 weeks of growth. However, the present study registered higher root/shoot ratios in mycorrhizal papaya at the end of 4 months of growth. This could be due to higher dry weight of mycorrhizal plants than control plants. de Silva et al. (1996) reported similar results in strawberry after 90 days of growth. The shift in root length and shoot dry weight ratios is a typical response to improved P nutrition in both mycorrhizal and non-mycorrhizal plants. In mycorrhizal plants, the P concentrations per unit dry weight are higher, thus, P use efficiency is lower than non-mycorrhizal plants (Marschner and Dell, 1994). This indicates that other growth factors (carbohydrates, mineral nutrients ) become limiting or that in mycorrhizal root systems the feed-back regulation between nutrient uptake rates and shoot demand are less well-regulated than in non-mycorrhizal plant systems (Douds et al., 1988). ACKNOWLEDGEMENTS Shri. Waman M. Khade Ex-director of Agriculture Department, and the Directorate of Agriculture State Government of Goa are thanked for assistance to carry out research work. REFERENCES
Copyright © 2009, African Crop Science Society The following images related to this document are available:Photo images[cs09015t3.jpg] [cs09015f3.jpg] [cs09015f1.jpg] [cs09015t2.jpg] [cs09015t1.jpg] [cs09015f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}