 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 18, No. 3, 2010, pp. 141 - 146 Short Communication INTROGRESSION OF STAY-GREEN TRAIT INTO A KENYAN FARMER PREFERED SORGHUM VARIETY K. NGUGI, W. KIMANI1 and D. KIAMBI1 University of Nairobi, Faculty of Agriculture, Department of
Plant Sciences & Crop Protection,
P. O. Box 29053 00625, Nairobi, Kenya
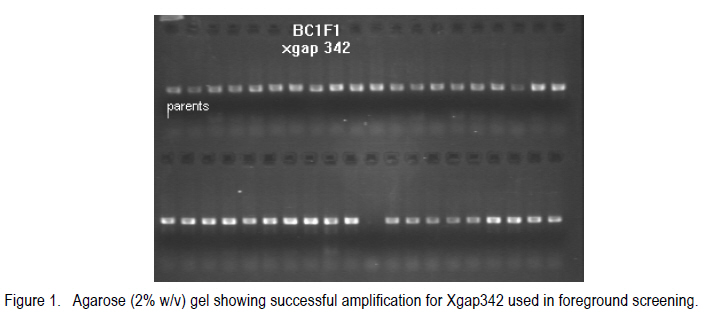
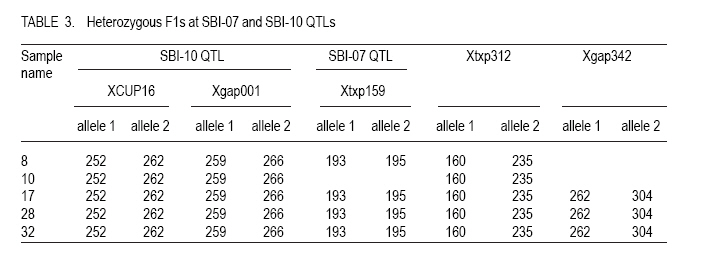
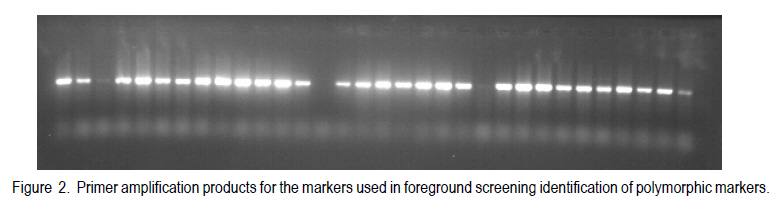
Corresponding author: kahiu.ngugi@yahoo.com; williskim@yahoo.com Code Number: cs10017 Abstract Backcross breeding enables breeders to transfer a desired trait from a Genetic Improvement of Kenyan sorghum variety for drought resistance donor parent, into the favoured genetic background of a recurrent parent. This study utilised back-cross breeding to transfer stay green quantitative trait locus (QTLs) from the donor parental line E36-1 into a Kenyan farmer-preferred variety, Ochuti as the recurrent parental line. The parental lines E36-1 has 3 stay green QTLs, SBI-01, SBI-07 and SBI-10 located at various chromosomes. The transfer of these QTLs was confirmed with the help of Simple Sequence Repeats (SSRs) molecular markers. Five foreground markers that were polymorphic among the two parental genotypes were used to identify individuals of F1 generation that had stay green QTLs transferred into Ochuti. A maximum of two QTLs, namely, SBI-07 and SBI-10 were identified as having been transferred into three individual genotypes. Two other F1 genotypes had only one QTL (SBI-10) transferred into Ochuti. The heterozygous F1 genotypes were used as the female parents in the generation of BC1F1. About 25% of the BC1F1 progenies that were genotyped had at least One QTL introgressed. As is the case in all marker-assisted back-cross breeding, the rate of success in introgressing QTL from donor to recurrent parental lines depends on the number of plants screened. Key Words: Back-crossing, molecular makers, Sorghum bicolor RÉSUMÉ L'améloration par croisement en retour permet les amélirateurs à transférer le trait désiré d'une variété de sorgho Kenyan génétiquement amélioré pour parent donneur de résistance à la sécheresse, en parent récurrent d'un passé génétiquement favori. Cette étude a utilisé le croisement en retour pour transférer le trait quantitatif locus de la persistance chlorophyllienne (QTLs) d'une lignée de parent donneur E36-1 dans la variété Kenyan préférée par les fermiers, Ochuti comme lignée parentale récurrente. La lignée parentale E36-1 a 3 QTL de persistence chlorophyllienne SBI-01, SBI-07 et SBI-10 localisés sur divers chromosomes. Le transfert de ces QTL était confirmé avec l'aide des marqueurs moléculaires à répétition simple de séquences (RSS). Cinq marqueurs rapprochés qui étaient polymorphoques parmi les deux génotypes parentaux étaient utilizes pour identifier les individus de génération F1 qui avaient le QTL de persistence chlorophyllienne transférée dans Ochuti. Un maximum de deux QTLs appelés SBI-07 et SBI-10 étaient identifié comme ayant été transférés dans trois génotypes individuels. Deux autres génotypes F1 avaient un QTL (SBI-10) transféré dans Ochuti. Les génotypes héterozygotes F1étaient utilisés comme de parents femelles dans la génération de BC1F1. Environ 25 % de progénies BC1F1 qui étaient génotypés avaient au moins un QTL introgressé. Comme c'est le cas dans toute amelioration par croisement de retour avec marqueur assisté, le taux de réussite en introgressant le QTL du donneur aux lignées parantales récurrentes dépend du nombre de plants testés. Mots Cles: Croisement de retour, marqueurs moléculaires, Sorghum bicolor Introduction Sorghum (Sorghum bicolor L.) is one of the most important cereals in the world and is ranked fifth according to the FAOSTAT data (FAOSTAT, 2006). It is a staple food for millions of people in the semi-arid tropics of Africa and Asia. Sorghum probably originated in Ethiopia and has spread to other parts of Africa, India, Southeast Asia, Australia and the United States (Dogget and Rao, 1995). Grain sorghum yields are very low in eastern Africa compared to the USA and well below the genetic potential. The national mean yields range from 0.6 to 1.5 t ha-1 as compared to 4.3 Mg ha-1 in the US (FAOSTAT, 2006). The biotic and abiotic production constraints to yield are numerous with combined effects of high yield losses. They include: soil water deficits, stalk borers, nitrogen deficiency, Striga sp. and other weeds, shoot fly, phosphorus deficiency, rust, smut, anthracnose, ergot and bacterial streak. Soil water deficits and Striga sp. are the most important constraints in Kenyan sorghum production (Wortmann et al., 2007). Drought in sorghum can occur before flowering (pre-flowering drought) or after flowering (post-flowering). Post-flowering drought stress is prevalent in many of these regions and this greatly affects the yields of sorghum due to premature leaf and plant senescence, stalk lodging and charcoal rot, and a reduction in seed size. However, some sorghum genotypes show tolerance to post-flowering drought (Rosenow, 1983; Rosenow and Clark, 1995). These genotypes have been referred to as `stay-green' types because they retain chlorophyll in their leaves and retain the ability photosynthesize even during post-flowering drought (Borrell et al., 2001). Stay green QTLs have been mapped in a number of genotypes such as E36-1, KS19, SC56 and B35 (Xu et al., 2000). Genotypes with the stay green trait take up more nitrogen from the soil and this increases the specific leaf nitrogen. This delays the onset and reduces the rate of senescence. The ultimate results are reduced lodging, resistance to charcoal rot, and proper grain filling even under water limiting conditions. The work reported here sought to introgress stay green QTL from an ICRISAT sorghum line E-36 well-characterised for stay-green QTL, into the background of the Kenyan farmers' preferred sorghum variety, Ochuti. MATERIALS AND METHODS F1 and BC1F1 generation. Seeds of inbred lines of Ochuti (Recurrent Parent) and E36-1(Donor Parent) were sown in a greenhouse at the College of Agriculture and Veterinary Sciences (CAVS), University of Nairobi in May 2009 and two leaves of each parent harvested 14 days after sowing. The leaves were preserved in 70% ethanol in Eppedorf tubes and then transferred to Biosciences East and Central Africa (BECA) laboratories at the International Livestock Research Institute (ILRI) Nairobi for genotyping. At flowering, artificial hybridisation was done by hand emasculation and pollination. Normal bisexual florets of Ochuti were hand emasculated and pollen from E36-1 transferred to the stigma of emasculated florets. The resulting F1 seeds were sown and the leaves of 96 F1 progenies harvested and stored in 70% ethanol in eppendorf tubes for genotyping to identify heterozygozity The heterozygous individuals were back-crossed to Ochuti (Recurrent parent) to generate BC1F1 genotypes. A total of 128 individuals from this back-cross were sown and their DNA isolated for genotyping to identify the individuals with the stay green QTLs (foreground screening) as well as those identify with the highest proportion of the recurrent parent. DNA extraction. The leaf from each plant was sampled individually and DNA extracted using the Cetyl-trimethyl Ammonium Bromide (CTAB) mini-prep method as developed by Mace et al. (2004). Two steel beads were inserted in each of the wells of strip tubes (Greentree Scientific, USA) of a Geno Grinder 2000, together with leaf samples and processed in liquid nitrogen. Pre-heated at 65 oC , 450µl of extraction buffer containing 3% (w/v) CTAB, 1.4M NaCl, 0.2 % (v/v) â-Mercapto-ethanol and 20 mM EDTA was added to the leaf samples and ground using the Genogrinder. The macerated substance was incubated for 15 minutes at 65 oC with occasional mixing. Solvent extraction was done by adding 450 µl Chloroform: isoamyalcohol (24:1) to each sample and mixed thoroughly by inversion. The tubes were centrifuged at 12000 rpm for 10 minutes at 24 oC and the upper portion transferred into fresh tubes (about 400 µl). Iso-propanol (0.7 volume) was added and inverted once to mix and the tubes were centrifuged after 20-30 minutes at 12000 rpm for 15 minutes. The supernatant was decanted and the pellet air dried for 30 minutes. A 200 µl low salt TE buffer (1 mM Tris and 0.1 m-1 M EDTA [pH 8]) with 3 µl RNase A (10 mg ml-1) was added to each sample and incubated at 37 oC in a water bath to remove the RNA. A second solvent extraction was done by adding 200 µl chloroform: isoamyalcohol (24:1) to each tube and inverting twice to mix and centrifuged. Later, the aqueous layer was transferred into fresh tubes. DNA purification was done by adding 315 µl ethanol and 1/10 volume of 3 M sodium acetate solution (pH 5.2) to each sample and then placing the samples in -20 oC for 5 minutes to allow precipitation. The tubes were then centrifuged at 12000 rpm for 5 minutes and the supernatant decanted. To wash the DNA pellet, 200 µl of 70% ethanol was added and centrifuged at 3500 rpm for 5 minutes. DNA pellet was air-dried for one hour and then re-suspended in 100 µl low salt TE [10 mM Tris, 1mM EDTA (pH 8)] buffer and stored at 4 oC. DNA Quality and purity check. DNA quality check was done using 0.8% (w/v) agarose gel electrophoresis stained with Gel red (5 µl 100 µl-1). The DNA quantity was checked using a nanodrop spectrophotometer. The required concentration for PCR reaction was 10 ng ml-1. Identification of polymorphism in foreground markers. A set of 24 foreground SSR markers (Table 1) was used for genotyping. The SSR markers used were m-13 tailed primers and were selected from those that flank the QTLs that confer stay-green trait in E36-1 sorghum variety. The forward primers were labelled with fluorescent dyes; FAM, NED, VIC and PET (PE-Applied Biosystems) since the amplicons were to be separated using a capillary electrophoresis. The PCR components for a 10 ml reaction were: 2 mM MgCl2, 0.20 ìM reverse primer, 0.04 ìM forward primer, 0.04 mM of each of the four dNTPs and 0.2 U AmpliTaq Gold DNA polymerase (AmpliTaq® with GeneAmp® Taq DNA polymerase: Applied Biosystems), 30 ng template DNA and top up to 10 ml reaction volume, double distilled water was added. Temperature cycling was carried out using the GeneAmp PCR systems 9600 (PE-Applied Biosystems) with the following protocol: 15 minutes at 94 °C , 40 cycles of 1 minute at 94 °C, 1 minute at 50°C and 2 minutes at 72 °C, with a final extension of 20 minutes at 72 °C. The PCR products were run on 2% (w/v) agarose gel electrophoresis to check the amplifications. Polymorphic molecular markers between two parental genotypes (ochuti and E36-1) were used to select true F1 by analyzing co-dominant markers having the alleles (bands) of both parental genotypes. A total of 96 F1 progenies were genotyped with the 7 polymorphic foreground markers between the two parents. Foreground screening for heterozygous BC1F1. A total of 128 BC1F1 DNA samples together with the two positive controls (the two parents) were genotyped with the five foreground SSR markers. The DNA that was extracted was amplified using PCR (Fig. 1) and the amplicons separated through capillary electrophoresis before allele calling on Gene Mapper software (V 3.7). Capillary electrophoresis. Genotyping was carried out by capillary electrophoresis using the ABI PRISM 3730 (Applied Biosystems). The loaded PCR products for capillary electrophoresis were mixed with 7.84 ml formamide (PE-Applied biosystems) and 0.16 ml GeneScan Liz 500 internal molecular weight size standard (orange) (Applied Biosystems). The DNA fragments were denatured at 95 oC for 3 minutes and then size fractioned using capillary electrophoresis. This system has automated sample loading and rapid electrophoresis (Buhariwalla and Crouch, 2004). Data analysis. The peaks were sized and the alleles called using the GeneMapper version 4.0 software and the internal GeneScan Liz 500 size standard. To ensure consistent results in each PCR and each capillary electrophoresis run, the positive control samples of the two parents were included in every electrophoresis run. RESULTS DNA quality and quantity check. Most of the samples had high molecular weight ranging from 16.45ng ml-1 in sample ID no.89 to 663.26 ng ml-1 in sample ID no.50. The optical density (OD) ratio (260 nm 280 nm-1) which helps assess the DNA quality ranged between 1.79 to 2.04 which indicates that the quality of DNA was ideal for PCR reactions. The pure DNA OD ratio 260 nm 280 nm-1 range is 1.8-2.0. Identification of heterozygous F1s. Out of the 96 F1 genotypes only five genotypes that had at least one stay green QTL introgressed. Genotypes ID no. 17, 28 and 32 had SBI-07 and SBI-10 QTLs introgressed, while genotype ID no. 8 and 10 had SBI-10 QTL introgressed (Table 3). SBI-01 QTL was not captured in any of the 96 genotypes. Foreground screening of BC1F1. Agarose (2% w/v) exhibited successful amplification (Fig. 1) and from a total of 128 individuals, 32 were found to be possessing one or two stay green QTLs after genotyping with the foreground SSR markers as shown in Table 3. Primer amplification products for the markers used in foreground screening are as shown in Figure 2. The 32 samples were progenies of genotypes ID no. 17, 28 and 32 (Data not shown). DISCUSSION DNA quality and quantity. The quality of DNA was appropriate for running PCR with SSR markers. The OD ratio (260 nm 280 nm-1) ranged from 1.8-2.0 which is an indication of good quality DNA. On 0.8% (w/v) agarose gel the bands were clear with no smear which is an indication of non-degraded DNA. Identification of polymorphic foreground molecular markers. The targeted QTLs from the donor parent E36-1 were SBI-01, SBI-07 and SBI-10. The markers selected were those that flanked the QTLs on both sides and one marker at the centre. For these markers to be of help in foreground selection in the subsequent generations, they have to be polymorphic meaning that they should show different allelic sizes between the two parents used in the cross (Collard et al., 2005). However most of the markers used were not polymorphic between the two parents making it difficult to discriminate between the parental lines. Foreground genotyping of F1 progenies. A total of five foreground markers were used to select heterozygous F1 genotypes and out of 96 individuals only 3 individuals that were heterozygous for the closest flanking markers in SBI-07 and SBI-10. Most of the other samples were homozygous for the female and the recurrent parent (Ochuti). This was an indication that that most of the individual genotypes were selfed and were not as a result of cross-pollination. Foreground genotyping of BC1F1. Two QTLs were possibly introgressed. The markers that are usually used for foreground screening should be tightly linked to the targeted QTL. The number of individuals screened for heterozygosity at the target loci determines the number of QTLs captured in every backcross generation. In this study, it was found that 22 genotypes in BC1F1 were sufficient to capture one QTL with a 95% (pd" 0.05) confidence. The number of individuals screened should increase with the number of back-crosses advanced, as there is an increase in recombination rate. CONCLUSION A maximum of two stay green QTLs were introgressed into Ochuti. The individual possessing these QTLs were used as one of the parents in BC1F1 generation. The large number of BC1F1 individuals used in this study enabled the identification of a number of genotypes with 1 or 2 stay green QTLs. Therefore, it is advisable for breeders to have at least 30 individual genotypes, in BC1F1 but should have a higher number than that in advanced generations in order to identify the targeted QTLs. References
Copyright 2010 - African Crop Science Journal The following images related to this document are available:Photo images[cs10017t3.jpg] [cs10017f3.jpg] [cs10017t1.jpg] [cs10017f1.jpg] [cs10017t2.jpg] [cs10017f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}