 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Crop Science Journal, Vol. 18, No. 3, 2010, pp. 147 - 154 COMBINING DROUGHT AND ALUMINIUM TOXICITY TOLERANCE TO IMPROVE SORGHUM PRODUCTIVITY T. Okiyo 1,2, S. Gudu 2, O. Kiplagat 2 and J. Owuoche 3 1Kenya Agricultural Research Institute, KARI Kibos,
P. O. Box 1490 - 40100, Kisumu, Kenya
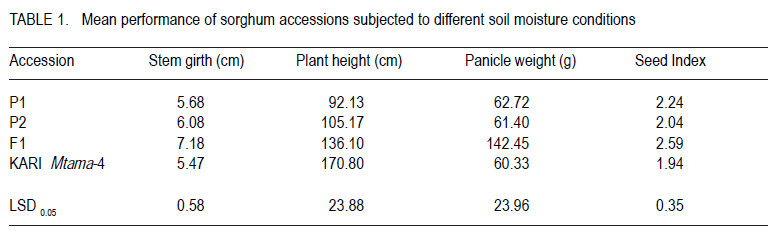
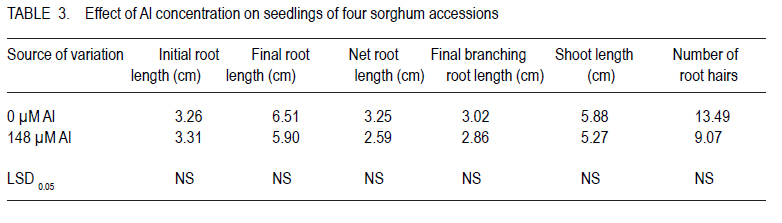
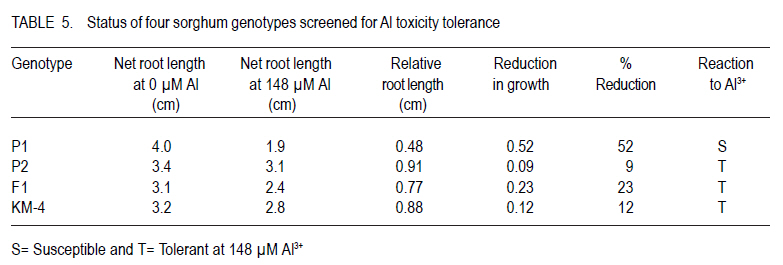
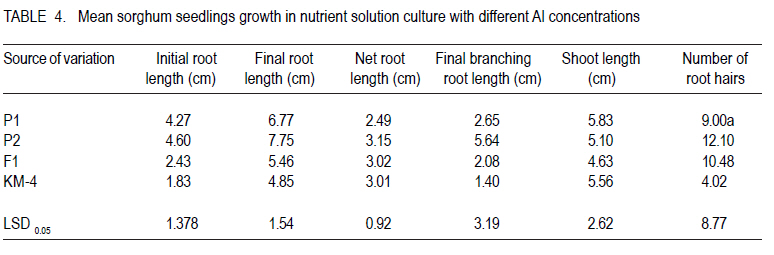
Corresponding author: tokiyo2006@yahoo.com Code Number: cs10018 ABSTRACT Moisture stress and aluminium toxicity in sorghum production can be overcome by breeding for tolerance. This study was set up to determine the response of sorghum (Sorghum bicolor L.) genotypes to post- anthesis drought and aluminium toxicity. Sorghum inbred P1 with stay green drought tolerance was crossed with P2, a standard aluminium tolerant cultivar. The parents, the first filial plants (F1) and KM-4 were tested for drought and aluminium toxicity tolerance in the field and solution culture with 0 and µMol 148 Al3+ respectively. The drought experiment was laid in RCBD with three replicates and split -plot arrangement. Randomly selected plots were denied irrigation from six weeks after emergence. There were significant (P < 0.05) differences between treatments and accessions. Panicle weight of F1 was outstanding and significantly different from the rest both with and without water. F1 out performed P1, P2 and KM-4 by 95.61, 146.37 and 328.81% without water; and by 155.44, 124. 27 and 82.02% with water, respectively. There were significant (P < 0.05) differences in the sorghum accessions with P2, KM-4 and F1 being tolerant. These results indicate that multiple stress tolerance can increase sorghum productivity. Key Words: Anthesis, Sorghum bicolor, stay green RÉSUMÉ Le stress hydrique et la toxicité aluminique dans la production de sorgho peuvent être surmontés par amélioration de la tolérance. Cette étude était faite dans le but de déterminer la réponse des génotypes du sorgho (Sorghum bicolor L.) à la sécheresse post-anthésis et la toxicité aluminique. Le sorgho de lignée pure P1 doté d'une persistence chlorophyllienne caractérisant sa tolérance à la sécheresse, était croisé avec P2, un cultivar de tolérance standard à l'aluminium. Les parents, les premières plantes filiales (F1) et KM-4 étaient testés au champs et avec une solution de culture 0 et 148 Al3+ µMol respectivement pour tolérance à la sécheresse et à la toxicité aluminique. Le dispositif experimental de l'essai sur la sécheresse était en blocs aléatoires complétement randomisés avec trois répétitions et un arrangement en split-plot. Les parcelles aléatoirement sélectionnées n'étaitent pas arrosées depuis six semaines après émergence. Des différences significatives (P < 0.05) étaient observées entre traitements et accessions. Le poids des panicules de F1 était exceptionnellement différent de celui avec ou sans eau. F1 avait été plus performant que P1, P2 et KM-4 avec 95.61, 146.37 et 328.81 % sans eau; et avec 155.44, 124. 27 et 82.02 % avec eau, respectivement. De différences significatives (P< 0.05) étaient aussi révélées entre les accessions de sorgho, le parent P2, les accessions KM-4 et les plants F1 étant tolérants. Ces résultats indiquent que la tolérance multiple au stress peut augmenter la productivité du sorgho. Mots Cles: Anthésis, Sorghum bicolor, persistance chlorophyllienne INTRODUCTION Drought is one of the most important abiotic stresses limiting sorghum (Sorghum bicolor) productivity around the world (Nguyen, 2008; Younesi and Moradi, 2009). It is certainly of great significance in the semi-arid tropics (SAT), where rainfall is generally low and its distribution is erratic (Ejeta at al., 1999; Annon, 2006), and yield losses of 70% do occur (Pocket, 2009). Arid and semi-arid lands (ASALs) cover 83% of Kenyan land mass (Kanyanjua et al., 2002). An effective and sustainable way to alleviate problems of crop production associated with drought is the development of crops that withstand moisture stress (Borrell et al., 2000; Nguyen, 2008). Photosynthesis per unit leaf area is decreased by water stress during the reproductive and grain filling periods (Dennis et al., 1984). Symptoms of post-flowering drought-stress susceptibility include premature leaf and plant senescence, stalk lodging and charcoal rot, and a reduction in seed size (Rosenow and Clark, 1995; Blum 2005). However, some annual cereals exhibit genetic variation for the degree or rate of leaf senescence during grain filling (Thomas and Smart, 1993). In sorghum, post-flowering drought tolerance is referred to as the stay green (Blum and Sulliva, 1986). This trait is exhibited by maintenance of green stems and upper leaves when water is limiting during grain filling (Rosenow, 1987). Sorghum genotypes with the stay-green trait continue to fill their grains normally even under limited water or moisture stress conditions (Borrell et al., 2000). Four genomic regions (Stg1, Stg2, Stg3 and Stg4) control the functional basis of `stay-green' in sorghum (Borrell et al., 2008). Haussmann et al. (2002) verified the general benefit of the stay-green trait for grain yield performance and stability. Work on sorghum germplasm with stay green in drought conditions by Borrell et al., (2001), confirmed that yield improvement of crosses with two sources of was directly by maintaining photosynthesis, and indirectly via improved lodging resistance. In acid soils with pH < 5.2, aluminium ions (Al3+) reduce root growth (Lidon and Barreiro, 2002) and hence, crop yield on potentially arable land by up to 50% (Bernel and Clark, 1998; Lang 2000). Al3+ was listed as a formidable phytotoxic barrier to crop production in acidic soils by (Vallagarcia et al., 2001). Acid soils cover some 40% of the earth's arable land (Giaveno et al., 2000; Gudu et al., 2001; Magalhaes et al., 2002) and 7.5% of Kenya`s land mass (Kanyanjua et al., 2002). A crop growing on Al3+ toxic soil suffers (phosphorus) deficiency despite sufficient levels. Low available delays flowering and leads to increased sensitivity to some foliar diseases further reducing production. Al3+ injury or toxicity in plants is manifested by inhibition of root growth (Peter et al., 1973; Gudu et al., 2001; Delhaize et al., 2004). Durieux et al. (1995) confirmed a decrease in the uptake of NO3 as a result of high aluminium levels in the soil; therefore, toxic effects of Al3+ aggravates soil infertility (Bushamuka and Zobel,1998). Sorghum growing on acid soils will yield less than their potential because of poorly developed roots that causes reduced nutrient and water uptake (Bernel and Clark, 1998; Anderson and Furlani, 2005). Soil amendments are very expensive, and if not carefully managed can destabilise the balance of natural ecosystem (Magalhaes et al., 2004). The cycle of soil fertility degradation and unsustainable agricultural production in the marginal areas can only be mitigated by using a holistic approach of appropriate strategies of soil nutrient management practices and improved crop varieties (Batiaono et al., 2006). Thus, screening of crop germplasm and developing appropriate varieties with high nutrient use efficiency could lead high crop productivity (Kochian et al., 2006). Variability in sensitivity in the major crops to Al3+ stress and the evaluation of root elongation in nutrient solution has been useful in developing Al3+ tolerant varieties ( Magalhaes et al., 2004; Spehar, 2004). Field evaluation though necessary for determining Al3+ toxicity tolerance, is quite expensive, time consuming and is frequently affected by large coefficients of variation due to the spatial variability in these soils (Gomez-Urrea et al., 1996). Field techniques to select for Al3+ tolerance need to be repeated along crop seasons to minimise the effects of uncontrolled environmental factors (Oettler et al., 2000). Results got in laboratory screening for tolerance correlate with performance in the field. The objective of this study was to set up to determine the response of sorghum genotypes to post- anthesis drought and aluminium toxicity. MATERIALS AND METHODS Plant material. A standard stay-green sorghum line with post-flowering drought tolerance was used as parent one (P1). It is a BP2derivative of IS12555; a durra landrace cultivar from Ethiopia (Walulu et al., 1994). It has large dark green pigmented and waxy leaves (IBPGR and ICRISAT, 1993). The panicle is semi- lax with erect spikes. The grains are dirty white (IBPGR and ICRISAT, 1993); awned and bold. It can produce up to 2 tillers depending on soil fertility and moisture availability. The main stem grows to about 95 cm and is early maturing. P1 was used as the female parent. The male parent (P2) is an Al3+ tolerant standard material from ICRISAT, which has tan green leaves with moderate wax. The panicle is semi-compact with small sized chalky white seeds (IBPGR and ICRISAT, 1993). It has a mature height of about 105 cm. The F1 was the first filial generation of hybridisation between P1 and P2, has large dark green pigmented waxy leaves. The panicle is semi-compact with medium to bold sized light brown seeds. It can produce up to 3 tillers. F1 has a mature height of 135 cm and early maturity period. F2 were got by advancing F1 progenies through controlled pollination. Sorghum variety KARI Mtama-4 (KM-4) was used in these studies as local check (LC) or farmer preferred variety. KM-4 was developed to suit medium rainfall conditions in western Kenya, and is, therefore, less adapted to drought. It is medium maturing, with grain yields of 4500 Kg Ha-1 and grows to an average height of 180 cm. The seeds are small and brown. The panicle has loose drooping primary branches. The response of KM-4 to Al3+ toxicity is not known. Field experiment for drought tolerance. Sowing was done by hand at seed rate of 8 kg ha-1. Staggered planting was done on 29th October 2008 for the crop to experience moisture stress after anthesis. The experiment was laid in randomised complete block design, with three replicates and split plot arrangement Three weeks after emergence the seedlings were thinned back to achieve a spacing of 50 cm x 30 cm. Single Super Phosphate was applied at planting in order to supply an equivalent of 40 kg P205. ha-1. A top dress of N at 45 kg ha-1 using urea in two splits of 20 and 25 kg was applied 30 days after emergence and at booting, respectively. Water treatment was considered the main-factor and genotypes as sub-factor. Although the crop was rain fed, a randomly selected main plot from each replicate received supplementary irrigation. The field was irrigated with about 25 mm every ten days for three sessions. Sorghum shoot fly, Atherigona soccata was controlled at early seedling stage and sorghum midge Contariana sorghicola at flowering using one litre of dimethoate ha-1. From a sample of 5 plants, stem girth, leaf width, flag leaf length, plant height and leaf colour were determined. Plant height was measured at harvest from the base of the plant above ground to the tip of the panicle. Days to 50% flower was recorded per plot. At maturity, panicles were harvested, dried, individually threshed and winnowed and 100 seeds counted. Weight of 100 seeds was recorded as seed index. Aluminium toxicity tolerance experiment in solution culture. Seeds were placed in distilled water to wet for 1 minute, then transferred to 0.1% NaOCl for eight minutes in order to sterilize. This was done in lamina flow chamber. The material was then rinsed eight times with distilled autoclaved water. The seed was then put in germinating dishes overlaid with filter paper moistened with sterilised water. The material was then placed in incubators at 25.1 oC and 66% RH (Relative Humidity), for three days (72 hours) in the dark. After 72 hours, initial root length (IRL) was measured. The seminal roots of the seedlings were inserted through the mesh bottoms of polyethylene cups, covered with black styrofoams beads before being placed into holes in the lids of polyethylene containers filled 8 liters of nutrient solution (49 seedlings per container). The basins were placed in growth chamber under continuous aeration with 26 oC day and 23 oC night temperatures, a light intensity of 550 µmol photons m-2 sec-1 and 12 hour photoperiod. The material was allowed 24 hours acclimation (Magalhaes et al., 2004); after which Al3+ treatment was administered. There were seven replicates in total. One replicate consisted of two basins, one control with zero Al3+ and the other with 148 µMol Al3+. The nutrient solution's pH was monitored daily and maintained at 4.2 by adding drops of HCl or Ca(OH)2 as appropriate. After 5 days, final root length (FRL), final root branching length (FRBL), shoot length (SL) and number of root hairs (NRH) were recorded. Intrinsic root growth rates were assessed for each individual using the root growth data obtained during the 5 days growth in the control solution. Accordingly, control growth rate (clgr 1d) was obtained as {clgr = FRL-IRL}. The root growth under Al3+ exposure over 5 days (Algr 5d), was calculated as, Algr5d = flAl flc, relative root length was calculated as RRL = NRL at 148 µM Al3+ over NRL at 0 µM Al3+, (Magalhaes et al., 2004). RRL was used as an indicator of growth reduction due to Al3+ toxicity and to determine the tolerance status of the sorghum accessions. The sorghum genotypes were classified as tolerant for 0- 20% reduction, moderately tolerant for 21-50% and susceptible for over 50% growth reduction. Data was analysed using SAS PROC GLM 1999 and means separated using LSD0.05. RESULTS Field experiment for drought tolerance. There were significant (P<0.05) differences in stem girth among accessions (Table 1). F1 had stems that were 23.82, 20.89 and 15.32% thicker than KM-4, P1 and P2, respectively. P2 and KM-4 were significantly different (P<0.05). KM-4 had the smallest stem girth, and F1 had the largest stem girth. Plant height was significantly different, with KM-4 having the tallest (170.8 cm) and P1 the shortest (92.13 cm). Overall, however, P2 with 105.17.cm was not significantly different from P1. F1 had a mean height of 136.1 cm; which was significantly different from the parents and the local check. P1, the genotype, was about ½ the height of KM-4, while F1 and P2 were 20.32 and 38.43% shorter, respectively. There was no significant difference in panicle weight of P1, P2 and KM-4. However, panicle weight of F1 was outstanding and significantly different from the rest both with and without water. F1 out performed P1, P2 and KARI Mtama-4 by 95.6, 146.37 and 328.81% without water; and by 155.44, 124.27 and 82.02% with water, respectively. P1 and F1 maintained green leaves upto harvest. F1 had the best overall performance, out yielding P1, P2 and KM-4 by 127.12%, 132.00% and 136.12%. F1 and P1 had bold seeds with an index of 2.59 and 2.24 while KARI Mtama-4 and P2 had small and medium sized seeds with SI of 1.94 and 2.04 (Table 1). Maturity period ranged from 61.05 days (watered) to 61.14 no water. Stem girth was 5.90 without water and 6.25 cm with water resulting in no significant (P>0.05) differences between the two treatments (Table 2). Panicle weight was 64.36 without water and 99.29 g with watere. Also, plant height was 114.08 and 138.00 cm without water and with water, respectively. These traits had significant differences (P < 0.05). B35 had the least yield reduction of 11.80% KARI Mtama-4 had the highest yield reduction of 69.96%. KARI Mtama-4 showed good yield 92.9 g and was second to F1 with 169 g on the watered plots but was the worst 27 g without water. Aluminium toxicity tolerance experiment in solution culture. The following traits, initial root length (IRL), final root length (FRL), were not significantly P>0.05 different at both 0 and 148 µ M Al3+ (Table 3 ). Net root length (NRL) and number of root hairs NRH were significantly different for the two treatments. Longer NRL and more root hairs were recorded at zero levels of Al3+ than at 148 µM (Table 3). Net root length was over 1 cm longer in the control than at 148µ M Al3+. There was no significant (P>0.05) difference effect on shoot length (Table 3 ). Net root length of F1 in nutrient culture at 148 µM Al3+ was 95% that of C1, 121% of B35 and 100 % or similar to that of KARI mtama-4 (Table 5). B35, the female parent (P1) and C1 the male parent (P2) had no significant differences (P> 0.05) in initial root elongation. They were, however, different from F1 and KM-4, yet the latter two were not significantly different (Table 4). Final root length was longest in C1 (7.75 cm), followed by B35 with 6.77, F1 5.46 and lastly with KM- 4 with 4.85 cm. Final branching root length had significant differences among the genotypes, yet B35 and C1 had no significant differences in FBRL but were quiet different from KM- 4 and F1. C1 however had the longest branching roots of 5.64 almost twice that of B35 and F1. The net root length had no significant difference (P>0.05), for C1, F1 and KM- 4. B35 had the shortest NRL that was significantly (P< 0.05) different from the rest (Table 4). B35 had the shortest mean (2.49 cm) (Table 4). There was no significant different in shoot length and number of root hairs among the sorghum accessions. However, C1 had more root hairs 12.10, followed by F1 with 10.48, while B35 had 9 and KM-4 had the least. C1 had the longest roots at 148 µM Al3+ (Table 4). There were distinct differences in the sorghum accessions response to Al3+ toxicity. B35 was most sensitive, with growth reduction of over 50%. KM-4, F1 and C1 were tolerant (Table 5) . The reduction in F1 was more than twice that of C1, the Al3+ tolerant parent. DISCUSSION Drought tolerance. That KARI Mtama-4 was second to F1 with water and the worst without water has a strong bearing on its drought tolerance and water use efficiency WUE. This variety was developed for medium rainfall areas of western Kenya .The variety also had the least stem girth and this tended to result in lodged stems further reducing mean yield. B35 suffered from red stem rot, caused by Colletotrichum graminicola, which compromised its performance. Genotypes possessing the stay-green trait maintain more photosynthetically active leaves than genotypes not possessing the trait. Stay-green genes delay drying of leaves, help the normal grain filling and reduce the incidence of lodging. This study confirms that choice of genotype is important for sustainable sorghum production in water deficient environments. This is because the F1 were the best at with or without water. The problem of dwindling water reservoirs and failing rains in SAT will definitely be relieved by developing high yielding and more adopted cultivars. Aluminium toxicity tolerance. There was reduction in root growth of all the genotypes tested at toxic level of Al3+ (Table 3). While working on sorghum Ohki (2006), noticed root growth inhibition as the most striking consequence of Al3+ toxicity. Common reaction in susceptible genotypes is poor plant growth as a result of root injury ( Anderson and Furlani, 2005; Jun ping et al., 2006). This is because root development plays a major role in a plant's response to water and nutrient availability. Poorly developed roots, therefore, lead to lesser exploration of bulk soil, hence, reducing nutrient and water uptake. Genotypes had different relative root length reductions at toxic Al3+ concentrations (Table 3). This shows variability in sensitivity to levels of Al3+ among the genotypes. Relative growth reduction in C1, F1 and KM-4 was significantly different though all had tolerant levels. Final branching root length was longest in the tolerant genotype C1. It had twice as much branching length at 148 µM Al3+ as P1 and F1. The local check had the shortest root branching length. Net root length was longest in P2, showing its ability to withstand toxic Al3+ concentration. (Table 2). The performance of the F1 was intermediate between the parents, but was only 0.13 cm shorter than the tolerant parent. P1, the drought tolerant but Al3+ toxicity susceptible parent had the least net root length. Though there were no significant differences in the number of root hairs, the tolerant parent had more hairs. F1 was intermediate between the parents. Shoot length varied significantly (Pd" 0.05) among the genotypes but did not respond to Al3+ treatments. This could be because the seedlings were still dependent on seed food reserves and not nutrients extracted from the growth media. ACKNOWLEDGEMENT This research was partially funded by Bio-Earn Sorghum Project awarded to Professor Samuel Gudu. I appreciate the Centre Director, KARI Kibos for allowing me to use Centre farm for experimentation. REFERENCES
Copyright 2010 - African Crop Science Journal The following images related to this document are available:Photo images[cs10018t3.jpg] [cs10018t2.jpg] [cs10018t5.jpg] [cs10018t4.jpg] [cs10018t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}