 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
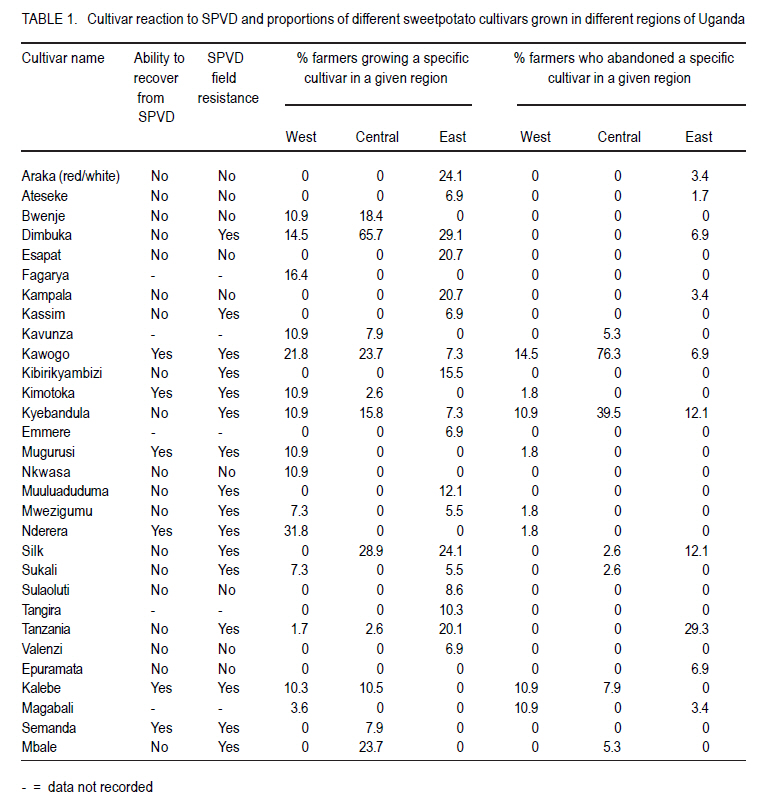
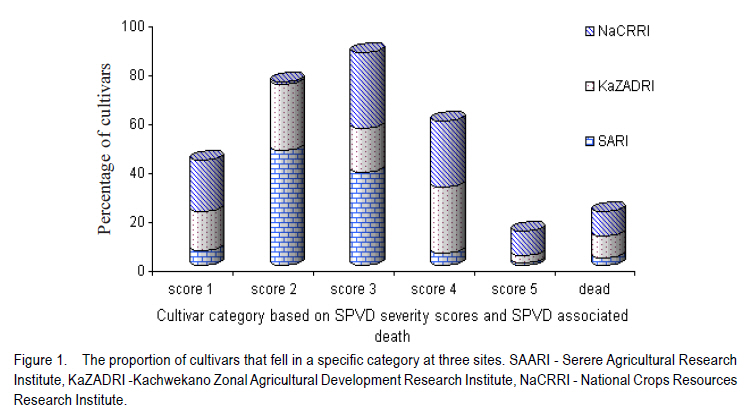
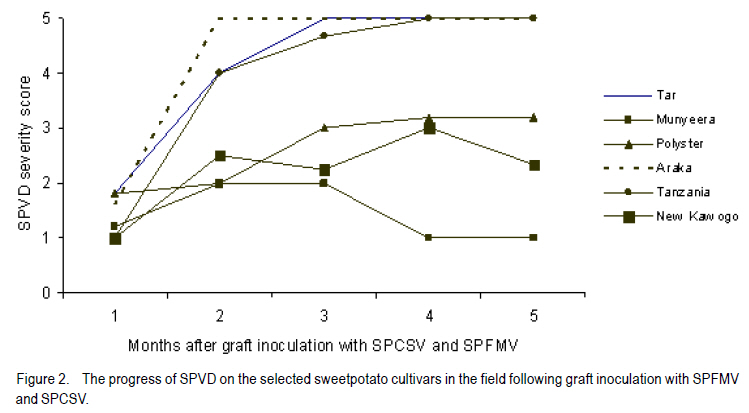
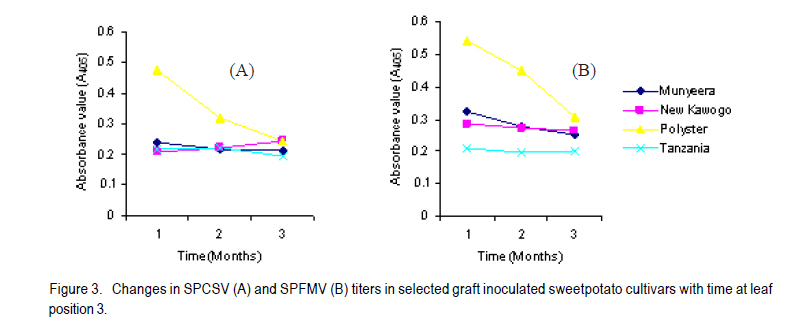
African Crop Science Journal, Vol. 18, No. 3, 2010, pp. 195 - 205 PREVALENCE AND IMPLICATIONS OF SWEETPOTATO RECOVERY FROM SWEET POTATO VIRUS DISEASE IN UGANDA E. Gasura and S.B. Mukasa1 Department of Crop Science, University of Zimbabwe, P. O. Box
MP167, Mount, Pleasant,
Harare, Zimbabwe
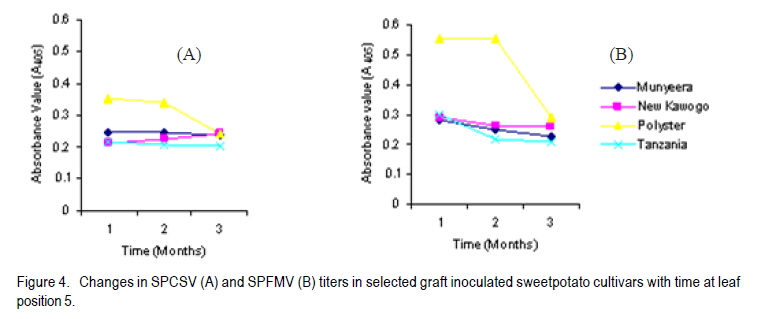
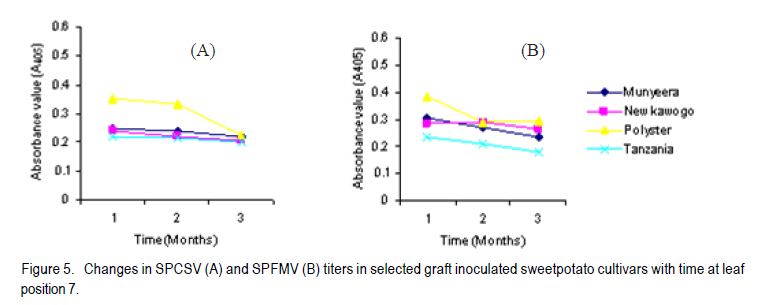
Corresponding author: sbmukasa@agric.mak.ac.ug Code Number: cs10023 ABSTRACT Sweet potato virus disease (SPVD) is the most important disease of sweetpotato in the tropics. It causes yield losses of up to 98% and extinction of elite cultivars. Although there are no reports of immune cultivars, disease recovery phenomenon (a manifestation of some form of resistance) was reported in many vegetatively propagated crops including sweetpotato. A total of 1320 sweetpotato cultivars from 3 major sweetpotato growing regions in Uganda were field evaluated for SPVD severity, of which 24% showed recovery from SPVD. The nature of SPVD recovery was assessed in the screenhouse using 20 distinct cultivars after graft inoculation with SPCSV and SPFMV, the SPVD causing viruses. SPVD progress in 20 graft inoculated cultivars was significantly (P<0.05) different; Munyeera was the most resistant cultivar followed by New Kawogo and Polyster. The 3 resistant cultivars (disease score 1-2) displayed recovery from SPVD which was correlated with a reduction in SPFMV titers as observed from quantitative ELISA. Reversion was not evident since all cuttings taken from recovered vine tips later tested positive for both viruses. Cultivars with the ability to recover were common in farmers' fields in central (36.4%) and western (33.3%) regions of Uganda, which are high SPVD pressure zones. Conversely, fewer cultivars (5.6%) from the low disease pressure eastern region displayed recovery. The very susceptible cultivars (disease score of 4-5) were not prevalent in central and western regions. These observations suggest that SPVD recovery has an influence on the cultivars grown in different SPVD pressure zones, where more susceptible cultivars are preferred in areas of low disease pressure. Key Words: Ipomoea batatas, resistance, reversion, SPCSV RÉSUMÉ La maladie de virus de la patate douce (SPVD) est une maladie la plus importante de la patate douce dans les tropiques. Elle provoque des pertes de production allant jusqu'à 98 % et voir même l'extinction des cultivars d'élite. Bien qu'il n'ya pas de rapports sur les cultivars résistants, le phénomène de récouvrement contre la maladie (manifestation de quelques formes de résistance) était signalé dans beaucoup de cultures végétativement propagées dont la patate douce. Au total 1320 cultivars de patate douce provenant de 3 grandes régions cultivatrices de la culture en Ouganda étaient évalués au champs pour la sévérité en SPVD parmi lesquels 24% ont montré une résistance à la SPVD. La nature de la résistance au SPVD était évaluée en serre utilisant 20 différents cultivars après inoculation de la greffe avec SPCSV et SPFMV, les SPVD causant les virus. Le progrès de la SPVD dans 20 greffes innoculées était significativement (P<0.05) différent; Munyeera était le cultivar le plus résistant suivi de New Kawogo et Polyster. Les 3 cultivars résistants (score de maladie: 1-2) ont montré un récouvrement de SPVD correlé avec une réduction dans les titres de SPFMV comme observé dans l'enzyme quantitatif ELISA. La réversion n'était pas évidente depuis que toutes les boutûres prises des cultivars résistants étaient finalement infectées par les virus SPCSVet SPFMV. Les cultivars ayant la capacité de se rétablir du virus étaient commun dans les champs de fermiers dans les régions centrale (36.4%) et ouest ( 33.3%) de l'Ouganda, qui étaient des zones sous pression élevée de SPVD. Au contraire, très peu de cultivars (5,6%) provenant de la région east à niveau bas de pression de maladie ont manifesté un rétablissement. Les cultivars les plus susceptibles (score de maladie: 4-5) n'étaient pas prévalents dans les régions centrale et ouest. Ces observations suggèrent que le récouvrement de la patate contre le SPVD a une influence sur les cultivars cultivés dans différentes zones de pression sous SPVD où plus de cultivars susceptibles sont préférés dans des milieux à basse pression de maladie. Mots Cles: Ipomoea batatas, résistance, réversion, SPCSV INTRODUCTION In Uganda, sweetpotato production is greatly constrained by the sweet potato virus disease (SPVD) that is caused by dual infection and synergistic interaction of an aphid transmitted, Sweet potato feathery mottle potyvirus (SPFMV) and a whitefly transmitted, Sweet potato chlorotic stunt crinivirus (SPCSV) (Gibson et al., 1998). SPVD is widespread in the major sweetpotato growing regions in Uganda and has been singled out as causing up to 98% yield loss and extinction of the early maturing and high yielding cultivars (Gibson et al., 1998). Single virus infections cause mild and transient symptoms or no symptoms at all in many sweetpotato cultivars (Mukasa et al., 2003). Co-infection of sweetpotato with SPCSV and SPFMV results in viral synergism leading to the development of severe symptoms termed sweet potato virus disease (SPVD), which includes stunting, leaf narrowing and distortion, chlorosis, mosaic or vein clearing (Gibson et al., 1998). Some plants are able to completely suppress virus multiplication, while others inhibit spread (Fraser, 1990) through cell-to-cell movement (Valkonen, 1994) or vascular movement (Carrington et al., 1996). Following host resistance induced virus localisation, the newly growing organs may be symptomless, a phenomenon known as recovery. Cuttings from such recovered portions may grow without virus infection and this is called reversion. Recovery and reversion are genotype dependant (Fondong et al., 2000). Following a bi-parental mating design between a resistant paternal cultivar Munyeera and four maternal parents, some of F1 progenies also displayed some levels of recovery suggesting that this phenomenon has a genetic basis (Gasura, 2008). Resistance to viruses in sweetpotato has been proposed to be associated with restricted virus movement (Nakashima et al., 1993). In East Africa, many sweetpotato cultivars seem to be quite resistant to single infections of SPFMV and SPCSV, showing mild symptoms from which they recover (Mwanga et al., 2002; Mukasa et al., 2006). However, recovery from the SPVD complex and its implications have hitherto not been documented. The aim of this study was to assess the occurrence, prevalence and the implications of sweetpotato recovery from SPVD on cultivar deployment and management of the disease. MATERIALS AND METHODS Occurrence and prevalence of SPVD recovery. A total of 1,302 sweetpotato cultivars were collected from 3 out of the 5 major agro-ecological zones of Uganda. Vines were sampled from 10 farmers' fields per district for 6 districts in each region, namely central, western, and eastern Uganda. The collection was assembled at the National Crops Resources Research Institute (NaCRRI), Namulonge, and used for the multi-site evaluation of the severity of sweetpotato virus disease (SPVD). The cultivars included locally bred clones, introductions and landraces. The severity evaluation experiments were conducted at Kachwekano Zonal Agricultural Development Research Institute (KaZADRI), NaCRRI and Serere Agricultural and Animal Research Institute (SAARI) that are located in western, central and eastern Uganda, respectively. SPVD severity was scored for each cultivar every month for four months at these three sites using a scale of 1-5, where 1 represents a healthy looking plant and 5 a severely diseased plant (Hahn, 1979). The number of plants which died from SPVD at each site was also recorded. The SPVD severity scores were subjected to analysis of variance (ANOVA) using GenStat software (GenStat, 2005) and the means were separated using the least significant difference (LSD) at 5% level of significance. For the three sites, a compound bar graph was plotted to show the relative proportions of cultivars out 1,302 that either fell into a given SPVD severity category or that died from SPVD. Based on the NaCRRI site, which had the highest SPVD incidence and severity, the cultivars were grouped into three categories. The first category showed the cultivars which died within the course of 4 months after infection with SPVD. The second and third groups were based on the severity scores. When the severity score increased progressively from low to high, the cultivars were termed non-recovering, and recovering when the severity score changed from high to low. Cultivar recovery from SPVD. From the 1,302 cultivars, 20 cultivars were chosen based on yield and quality attributes, and reaction to SPVD, and used in the recovery experiment following graft inoculation. The cultivars consisted of land races (Araka Red, Araka White, Bitambi, Kikuyu, Kisakyamaria, Kyebandula, Munyeera, Polyster, Silk, Sudan, Tanzania, Tar and Zapallo), improved varieties (Bwanjule, Enjumula, Kyebandula, NASPOT 1, New Kawogo and Sowola), and introductions (Beauregard and Nashu). The 20 healthy looking sweetpotato cultivars were potted and raised in the screenhouse at Makerere University Agricultural Research Institute, Kabanyolo (MUARIK). A leaf from cultivar Tanzania infected with a combination of SPFMV and SPCSV, which cause SPVD, was graft inoculated to each of the twenty cultivars. The graft inoculated plants were diagnosed using serology to confirm infection by SPCSV and SPFMV. Detection of SPFMV was carried out using nitrocellulose membrane enzyme-linked immunosorbent assay (NCM-ELISA) as described by Gibb and Podavan (1993). Also, SPCSV was diagnosed using the triple antibody sandwich ELISA (TAS-ELISA) as described (Gibson et al., 1998). The 20 graft inoculated cultivars were then transferred to the field at MUARIK and planted in a completely randomised design with 5 replications. Disease severity was scored once a month for six months, using a scale of 1-5. Where, 1 represented a healthy looking plant and 5 a severely diseased plant as described by Hahn (1979). Virus titers in resistant and susceptible cultivars. Based on the above study, the 20 sweetpotato cultivars were classified as resistant, moderately resistant or susceptible. Two resistant cultivars (Munyeera and New Kawogo), one moderately resistant cultivar (Polyster) and a susceptible control (cultivar Tanzania) were potted in a screenhouse. From each cultivar, three sample leaf discs of 1 cm diameter were taken from leaf positions number 3 (the first fully opened leaf from the shoot apex), 5 and 7 as described by Mukasa et al. (2006). Virus detection and quantification were done using DAS-ELISA (SPFMV) and TAS-ELISA (SPCSV) as described by Gibson et al. (1998). Using the GenStat software, ANOVA was carried out in a 2 (virus) X 3 (time) X 4 (cultivars) factorial option. Mean absorbance values were separated using LSD (5%) where significant differences were occurred. Fully recovered vine tips of less than 30 cm from the resistant sweetpotato cultivars (Munyeera, New Kawogo and Polyster) were collected and potted in 30 replications. Detection for both viruses using DAS-ELISA (SPFMV) and TAS-ELISA (SPCSV) was done and symptoms were recorded. Negative results were verified using RT-PCR. Total RNA was extracted from 200 mg of the leaf tissue using the TRIzol LS Reagent (Invitrogen Ltd) according to the manufacturer's instructions. It was quantified using a NanoDrop spectrophotometer (NanoDrop Technologies, Inc.). The concentration of the RNA was equalized and 1 mg used in cDNA systhesis as described (Mukasa et al., 2006). RT-PCR was done using a RevertAid First Strand cDNA Synthesis Kit (MBI Fermentas) with the following gene specific forward and reverse primers: CP1 (CTG CTA GAT TGT TAG AAA) and CP2 (TAT ATG AAA ATA TAG TTC) for SPCSV (Alicai et al., 1999) and PVD1 and Poly-T for SPFMV (Mukasa et al., 2003). RESULTS Sweetpotato cultivar composition. Out of the 1,302 sweetpotato accessions collected from the 3 major sweetpotato growing regions, 202 distict cultivars were identified in different farmer fields; 80, 66 and 56 cultivars were from the western, central and eastern study regions, respectively. Some cultivars had more than one name; for instance Dimbuka also called Bunduguza, Tanzania sometimes called Osukutu, and Esapat also called Boy or Osapat. The most popular name for each cultivar was chosen in this report. However, 15, 16 and 21 cultivars were found to be widely grown in the west, central and east, respectively (Table 1). Cultivars like Bwenje, Dimbuka, Kawogo, Kavunza, Kimotoka, Kyebandula and Kalebe were common in both the western and central regions that are high SPVD pressure zones. Most of these cultivars have shown either field resistance or the ability to recover from SPVD (Table 1). Cultivar Fagarya was found only in the western region. Cultivars Mbale, Semanda and NASPOT 1 were only found in the central, whereas cultivars Araka, Ateseke, Esapat, Kampala, Kibirikyambizi, Sulaoluti, and Muuluaduduma were only found in the eastern region. Cultivars Dimbuka, Kawogo, Kyebandula, Tanzania, and Sukali were widely grown in all the 3 regions. SPVD was observed in all farmers' fields in the central and western regions where the majority of resistant plants were found (Fig. 1). However, in the eastern region where most cultivars grown were susceptible to SPVD, most of the cultivars had low to no symptoms of SPVD. It was interesting to note that the cultivars reported as most commonly abandoned in all regions were Kawogo, Kyebandula and Dimbuka yet these were at the same time the most widely grown.This was partly because some cultivars abandoned by one farmer could be still found in production with the next farmer. Reseaons given by farmers for abandonment of some sweetpotato cultivars include poor yields, lack of quality planting material (susceptible to diseases), taste (low dry matter), late maturity, and susceptability to drought. Prevalence of SPVD recovery. Severe symptoms typical of SPVD were observed on many cultivars planted at different sites. Cultivar severity scores were significantly (P<0.05) different at four months after planting (Fig. 1). Furthermore, the site effect was significant, with NaCRRI site having the highest severity scores, followed by KaZADRI. SAARI had the lowest scores (Fig. 1). At SAARI, the majority of the cultivars had the severity score of 2 or 3. At KaZADRI, the cultivars were more in severity score 1-4 and relatively few in the severity score of 5. At NaCRRI, many cultivars were in the group 1, 3 and 4, with very few cultivars in the group 2 (Fig. 1). Cultivars that died were relatively many at NaCRRI (10%) and KaZADRI (9%) but low at SAARI (3%). Based on the NaCRRI site (which had the highest SPVD incidence and severity), these cultivars were grouped into two categories according to the severity scores. Out of the cultivars evaluated (1,302), 66% were grouped as non-recovering and 24% were grouped as recovering. However, some (10%) died after infection with SPVD during the period of four months and this death was associated with SPVD. Sweetpotato cultivar recovery from SPVD. Typical SPVD symptoms occurred on the study materials which were graft inoculated with SPFMV and SPCSV (Fig. 2). With the exception of the cultivar Munyeera, New Kawogo and Polyster, the rest of the cultivars succumbed to SPVD and failed to recover, instead their disease severity scores either remained constant or increased. Cultivars Munyeera and New Kawogo recovered from the mean severity scores of 2.3 and 3.5 to mean severity scores of 1.2 and 2.3, respectively. Upon reaching the mean severity score of 3.5, cultivar Polyster neither increased nor reduced its severity score. Based on the ability to recover and disease severity scores, two cultivars Munyeera and New Kawogo were grouped as resistant. Cultivar Polyster was moderately resistant and the rest ranged from moderately susceptible to susceptible. Virus titers and SPVD recovery. Virus titers were significantly different (P<0.05) for both time and cultivars tested (Figs. 3, 4, 5). In general, the virus titers decreased with plant age. Although the titers decreased with time in all cultivars, the decrease was highly significant in the resistant cultivars. Resistant plants had nil to very low virus titers compared to the susceptible control for both viruses at all leaf positions tested (Figs. 3, 4, 5). When a heavy viral load was introduced in the test plants through graft inoculation, the three resistant cultivars allowed the virus to spread but later reduced viral titers unlike the susceptible plants. However, the titers of SPFMV were significantly lower in resistant cultivars than the susceptible cultivar Tanzania. To the contrary, titers of SPCSV were not significantly different irrespective of level of resistance displayed by the plants. Reduction of SPFMV titers was accompanied by a reduction in symptom severity leading to symptom recovery in these resistant genotypes. DISCUSSION Sweetpotato virus disease (SPVD) has been reported to cause up to 98% yield loss (Gibson et al., 1998). Therefore, the high yield reported by farmers can refer to SPVD resistance, tolerance or ability of the cultivar to escape SPVD. The differences in composition of the cultivars found in the eastern region and those found in western and central regions, therefore, reflect the effects of SPVD. Cultivars like Araka, Esapat, Kampala and Tanzania, commonly grown in the eastern Uganda where SPVD pressure is low are generally susceptible to SPVD. These cultivars neither posses field resistance nor the ability to recover from SPVD (Table 1). However, cultivars like Kawogo, Nderera, Mugurusi and Dimbuka found in the west and central region, where SPVD pressure is high, either have the ability to recover from SPVD or have some level of field resistance to SPVD (Table 1). Furthermore, some of these cultivars like New Kawogo were also reported to be resistant to SPVD (Mwanga et al., 2002). The occurrence of symptoms of SPVD in west and central but few to none in the east, further support the fact that the eastern region has low SPVD pressure (Tugume et al., 2008), which in turn allows the production of susceptible cultivars. Interestingly, some highly susceptible cultivars like Silk and Kyebandula were also found in all regions despite the SPVD pressure level (Table 1). This suggests the influence of other factors than SPVD. Farmers reported to grow these cultivars mainly because of their taste and high dry matter content in addition to yield and early maturity. Farmers who abandoned the cultivars were found in both low and high SPVD pressure zones. Cultivar Tanzania had high levels of abandonment in the eastern region. Cultivar abandonment can, therefore, reflect a general loss in fitness of a cultivar overtime resulting in low yields. For example, cultivar New Kawogo though resistant to SPVD is low yielding. Despite, the loss in cultivar fitness, some farmers were often reluctant to discard their once elite cultivar. This could explain some varieties which are abandoned by some farmers could still be found with other nearby farmers. However, farmers in western region reported both low yield and disappearance of planting materials more often than those in the east. Cultivar abandonment in western region could be, therefore, exacerbated by high SPVD pressure. Furthermore, SPVD has been reported to cause up to 98% yield loss and causing the disappearance of some once elite cultivars (Gibson et al., 1998). The majority of SPVD resistant plants are also low yielding as evidenced by New Kawogo and Munyeera. This is in line with the idea that the early maturing and high yielding materials are disappearing. This calls for efforts to breed for SPVD resistance while maintaining the cultivar yield. The high SPVD severity at NaCRRI and KaZADRI (Fig. 1) indicates that these are high SPVD pressure zones. Mukasa et al. (2003) reported the highest incidences of SPFMV and SPCSV in central Uganda, followed by western region where NaCRRI and KaZADRI are located. This further supports the high level of SPVD pressure observed in this study. However, SAARI had the least SPVD severity scores. This probably explains why some cultivars like Araka and Tanzania which either had high severity scores at NaCRRI or died were able to survive at SAARI where they displayed low severity scores. Although many cultivars showed high SPVD symptom severity which was associated with death of some cultivars, some showed the ability to recover from SPVD under natural field infection (Table 1). This recovery is a natural phenomenon that can be attributed to a manifestation of some form of resistance. Recovery was once reported from single infections with SPFMV and SPCSV (Mukasa et al., 2006). However, some cultivars failed torecover from SPVD. Failure to recover from SPVD and the subsequent death of some cultivars is not surprising. For instance, it was reported that during dual infection and synergistic interactions, resistance to single infections is compromised (Kreuze et al., 2005) leading to elevated titer levels of SPFMV (Karyeija et al., 2000), thus causing SPVD (Gibson et al., 1998). Of the cultivars evaluated, the landrace Munyeera was the most resistant to SPVD, followed by New Kawogo and Polyster. New Kawogo was reported by Mwanga et al. (2002) to have average SPVD severity scores of 2.3 to 4.3. Thus, Munyeera could be used as a new source of modest resistance in preference to both New Kawogo and Polyster, unless the breeding programmes prefer some other traits not found in Munyeera. Cultivars Tanzania, NASPOT 1 and Sowola were reported to be resistant (Mwanga et al., 2002), but data from this study are not in agreement (data not shown for cultivars Sowola and NASPOT 1). The ability to recover from SPVD was observed in cultivar Munyeera, New Kawogo and Polyster following co-infection with SPCSV and SPFMV, unlike in single infections from which Mwanga et al. (2002) observed recovery. Mukasa et al. (2006) also found that in coinfections, recovery is compromised in cultivars Dimbuka and Tanzania which are resistant to single infections. The breaking down of extreme resistance to SPFMV following co-infection with SPCSV observed in most of the cultivars is not unusual. However, in this study the three resistant cultivars showed recovery from the SPVD complex showing their potential to be used as SPVD resistance sources. Gibson et al. (1998) noted that SPVD in its extreme case can lead to the extinction of some cultivars. Fraser (1990) reported that genetically controlled resistance is occurs when some individuals of the same species are able to resist pathogen attack, whereas some are susceptible. Accordingly, some cultivars could recover from SPVD when others succumb or die, suggesting that the recovering cultivars may have some level of resistance to this disease complex. In fact, Gasura et al. (2008) reported segregation of the recovery phenotype in sweetpotato. The three cultivars, Munyeera, New Kawogo and Polyster,which did not completely succumb to SPVD following graft inoculation seem to have some level of genetically controlled resistance. Most of the cuttings taken from the symptomless upper parts of the recovered vine tips of cultivar Munyeera and New Kawogo, and potted in the screen house, sprouted without symptoms and tested negative with serology. The negative ELISA results could partly explained by the low virus titers in leaves from recovered vine tips because the recovered vines later developed symptoms and tested positive with RT-PCR. This observation is consistent with previous reports showing that East African sweetpotato cultivars express resistance to single virus infections which is characterised by lack of symptoms and virus titers below the detection threshold level of serology (Mwanga et al., 2002). Shukla et al. (1994) observed the same in single infections and argued that serology has low detection limits, at least 1-10 ng of virus per kilogramme of infected tissue. Karyeija et al. (2000) and Mukasa et al. (2006) also reported that plants that tested negative from ELISA could test positive using sensitive methods like grafting on Ipomoea setosa plants. Reversion was not observed in sweetpotato unlike in cassava as reported by Storey and Nichols (1938) with respect to African cassava mosaic virus disease. This could suggest that resistance to SPVD is through titer reduction leading to symptom recovery. Obtaining planting materials from old fields has a danger of rampantly spreading different diseases such as SPVD. This is because such materials may have infections or latent infections of some diseases despite their healthy looking. However, farmers select healthy looking vines to use as planting materials because such materials normally give better yield. Gibson and Aritua (2002) also noted that farmers tend to rogue infected plants and also plant asymptomatic vines to alleviate yield loss from SPVD. This implies that most of the planting material in SPVD high pressure zones come from those cultivars which have the ability to recover such as New Kawogo, Nderera and Munyeera. Such cultivars with the ability to recover are generally low yielding. Over time, this practice of selecting recovered vines results in increased proportions of recovering cultivars. This can be supported by the differences in composition of cultivars found in the east and those of the west and central regions (Table 1). The cultivars with the ability to recover comprise 36.4, 33.3 and 5.6% of the popularly grown cultivars in the central, western and eastern regions, respectively. This shows that in the high SPVD pressure zones (central and western regions) the farmers unintentionally increase the proportion of the cultivars with the ability to recover. In the low SPVD pressure zone (eastern region) the farmers mainly grow cultivars which do not recover from SPVD since such cultivars also display high tuber yield. Incorporating recovery into the elite but susceptible cultivars could be very essential in enhancing resistance to SPVD and thus maintain or increase yield. The titers of SPFMV were low in all the leaf positions in the SPVD infected resistant plants. Further, they showed a gradual decline contrary to higher titers (600 fold increase at 5 weeks after inoculation) which should be found in dual and synergistic interactions (Karyeija et al., 2000). This clearly suggests that the synergism was somehow halted in resistant plants since SPCSV was found in lower leaves as expected in single or multiple infections whereas SPFMV was found in all leaf positions but in reduced titers (Figs. 3, 4). This suggests that resistance mechanism to SPVD is via overall in planta reduction in titers of the beneficiary virus (SPFMV), through reduced multiplication rather than movement in the plant. There are several reported examples of resistant host restricting cell to cell movement (Valkonen, 1994) or vascular movement (Carrington et al., 1996) of potyviruses. This was previously proposed as the possible resistance mechanism to SPFMV in sweetpotato (Nakashima et al., 1993) probably leading to recovery. Furthermore, Petty et al. (1990) reported that the level of viral accumulation affects its ability to move systemically. Our data did not support restricted virus movement as a mechanism for resistance to both SPFMV and SPCSV because these viruses were detected in both old and young leaves using RT-PCR even in the recovered vine tips. Furthermore, in graft inoculated plants, the potyvirus moved in resistant plants at the same rate as in susceptible plants as was evidenced by symptoms which first appeared within two weeks following graft inoculation in young leaves. This occurred almost at the same time, irrespective of the resistance level of the genotype suggesting that virus could still move throughout the plant. This suggests that the loading, unloading and distribution of SPFMV were not limited. ACKNOWLEDGEMENT We thank the National Crops Resources Research Institute (NaCRRI), Namulonge, Uganda for the provision of germplasm. Special thanks go to Dr. R.O.M. Mwanga and Ms. Agnes Alajo from NaCRRI for the assistance in experimental set up, monitoring and data collection. This work was funded by Rockefeller Foundation through the RU-CG005 grant of the RUFORUM programme and Sida/SAREC through the Sweetpotato project of the BIO-EARN programme. REFERENCES
Copyright 2010 - African Crop Science Journal The following images related to this document are available:Photo images[cs10023f3.jpg] [cs10023f1.jpg] [cs10023t1.jpg] [cs10023f2.jpg] [cs10023f4.jpg] [cs10023f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}